
Katekoliamiinien synteesi, erittyminen ja aineenvaihdunta
Viimeksi tarkistettu: 19.10.2021

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
Lisämunuaisten aivokerros tuottaa yhdisteen, joka on kaukana rakenteen steroideista. Ne sisältävät 3,4-dihydroksifenyyli (katekoli) ydin ja niitä kutsutaan katekoliamiineiksi. Näihin kuuluvat adrenaliini, norepinefriini ja dopamiini beeta-oksi- tiiniamiini.
Sekvenssi katekoliamiinipitoisuudet synteesi on varsin yksinkertainen: Tyrosiini → dihydroksifenyylialaniinia (DOPA) → → dopamiini noradrenaliinin → adrenaliinia. Tyrosiini tulee kehon ruoan kanssa, mutta voi olla muodostettu fenyylialaniinin maksassa vaikutuksen alaisena fenyylialaniinihydroksylaasin. Tyrosiinin transformaation lopulliset tuotteet kudoksissa ovat erilaiset. Lisämunuaisytimessä prosessi etenee vaiheeseen adrenaliini muodostumista, päissä sympaattisen hermoja - noradrenaliinin, tietyissä keskushermoston neuroneiden hermoston katekoliamiinien dopamiinin synteesiä täytetty lomake.
Tyrosiinin muuntaminen DOPA: hon katalysoi tyrosiinihydroksylaasi, jonka kofaktoreita ovat tetrahydrobiopteriini ja happi. Uskotaan, että tämä entsyymi rajoittaa katekoliamiinin biosynteesin koko prosessin nopeutta ja se inhiboi prosessin lopputuotteita. Tyrosiinihydroksylaasi on pääketarkoitus säätelyvaikutusten vaikutuksesta katekoliamiinien biosynteesiin.
Dopa muuntaminen katalysoi entsyymi dopa-dekarboksylaasin (kofaktorin - pyridoksaali), joka on suhteellisen ei-spesifinen ja dekarboksyloidaan, ja muita aromaattisia L-aminohappo. On kuitenkin olemassa viitteitä siitä, että mahdollisuus muuttaa synteesin katekoliamiinien muuttamalla aktiivisuus ja entsyymin. Joissakin neuroneissa puuttuu entsyymien muuntamiseksi edelleen dopamiinin ja että se on lopputuote. Muut kudokset sisältävät dopamiini-beeta-hydroksylaasin (kofaktoreita - kupari, askorbiinihappo ja happi), joka muuntaa dopamiinin ja noradrenaliinin. Lisämunuaisytimessä (mutta ei sympaattinen hermopäätteitä) on läsnä fenyylietanoliamiini - metyylitransferaasi muodostavan adrenaliini päässä noradrenaliinia. Metyyliryhmien luovuttaja tässä tapauksessa on S-adenosyylimetioniini.
On tärkeää muistaa, että synteesi fenyylietanoliamiini-N-Metiltransferazy glukokortikoidit, jotka kuuluvat aivokuoren kerroksen porttilaskimojärjestelmä. Tämä saattaa selittää se, valheita yhdistyy kaksi erilaista Umpieritysrauhaset yhdessä ruumiissa. Tarkoitettua glukokortikoidin synteesin adrenaliini korostaa se, että lisämunuaisytimessä tuottavien solujen norepinefriinin, jotka on järjestetty noin valtimoihin, kun taas verisolut saadaan adrenalinprodutsiruyuschie oleellisesti laskimoiden poskionteloiden, paikallinen lisämunuaisen kuorikerros.
Romahdus katekoliamiinien tapahtuu pääasiassa vaikutuksen alaisena kahden entsyymin järjestelmät: katekoli-O-metyylitransferaasin (COMT) ja monoamiinioksidaasin (MAO). Tärkeimmistä tavoista adrenaliinin ja noradrenaliinin rappeutuminen on esitetty kaavamaisesti kuviossa. 54. Toiminnan alaisena COMT läsnä ollessa luovuttajan metyyliryhmien S-adrenozilmetionina katekoliamiinien ja muunnetaan normetanefriini metanefriini (3-O-metyyli-johdannaiset adrenaliinin ja noradrenaliinin), joka vaikutuksen alaisena MAO muuntaa aldehydeiksi ja (n läsnä ollessa aldehydin) on vanillyyli-mantelihapon happo (ICH) - Päähajoamistuotteen noradrenaliinin ja adrenaliinin. Samassa asiassa, kun ensimmäinen altistetaan katekoliamiinien MAO toiminta, ei COMT, ne muunnetaan 3,4-dioksimindalevy aldehydi, ja sitten vaikutuksen alaisena aldehydin ja COMT - 3,4-dioksimindalnuyu hapon ja kierukka. Kun läsnä on alkoholidehydrogenaasi katekoliamiinien voivat muodostaa 3-metoksi-4-oksifenilglikol, tärkein lopputuotteen hajoamisen adrenaliinin ja noradrenaliinin keskushermostossa.
Hajoamisen dopamiini etenee samalla tavalla, paitsi että sen metaboliitit ovat vailla hydroksyyliryhmien beeta-hiiliatomiin, ja näin ollen sen sijaan, että vanillyyli-mantelihappoa muodostuu homo- (HVA) ja 3-metoksi-4-oksifeniluksusnaya happoa.
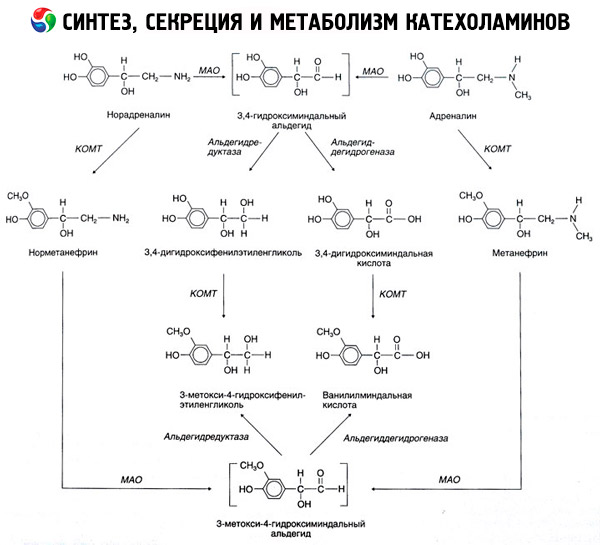
Voidaan myös olettaa, että kino- koliinien molekyylin hapettamista varten voidaan käyttää kino- kolyliamiinien hapettumista, josta välituotteilla on voimakasta biologista aktiivisuutta.
Muodostuu toiminnan sytosolientsyymien, adrenaliinia ja noradrenaliinia sympaattisen hermopäätteitä, lisämunuaisytimessä ja anna eriterakkuloissa joka suojaa niitä toiminnan hajottavien entsyymien. Katekoliamiinien keräytyminen rakeisiin vaatii energiakustannuksia. In kromaffiinisolujen rakeita lisämunuaisytimessä katekoliamiineihin sitoutunut tiukasti ATP: tä (suhteessa 4: 1) ja tiettyjen proteiinien - kromograniini, joka estää diffuusion hormonien rakeiden sytoplasmassa.
Suora ärsyke erittymiseen katekoliamiinien on ilmeisesti tunkeutuminen kalsium-solua stimuloiva eksosytoosia (fuusio kalvo rakeiden kanssa solun pinnalla ja niiden ero kokonaissaanto liukeneva - katekoliamiinien, dopamiini-beeta-hydroksylaasin, ATP: n ja Kromograniini - solunulkoiseen tilaan) .
Katekoliamiinien fysiologiset vaikutukset ja niiden toiminnan mekanismi
Katekoliamiinien vaikutukset alkavat vuorovaikutuksella kohdesolujen spesifisten reseptoreiden kanssa. Jos reseptorit kilpirauhasen ja steroidihormonien, sijaitsevat soluissa, katekoliamiinin reseptorit (sekä asetyylikoliinin ja peptidihormoneja) ovat läsnä solun ulkopinnan.
On pitkään todettu, että suhteen joidenkin reaktioiden adrenaliinia tai noradrenaliinia ovat tehokkaampia kuin synteettinen katekolamiini isoproterenol, kun taas toisissa vaikutus on parempi kuin toiminnan isoproterenolihoidon adrenaliinin tai noradrenaliinin. Perustuu tähän käsitteeseen läsnäolo kahden kudosten on kehitetty adrenergisen alfa ja beta, ja vain jokin näiden kahden voi olla läsnä joissakin niistä. Isoproterenolin on tehokkain agonistin beeta-adrenergisiin reseptoreihin, kun taas synteettinen yhdiste fenyyliefriinin - tehokkain agonisti alfa-adrenergisiin reseptoreihin. Luonnon katekoliamiinien - adrenaliini ja noradrenaliini - voivat olla vuorovaikutuksessa reseptorien kanssa molempien, mutta adrenaliini ottaa suurempi affiniteetti beta, ja norepinefriinin - alfa-reseptoreihin.
Katekoliamiinien vahvempi aktivoida sydämen beetareseptoreita kuin -beetareseptorien sileän lihaksen, jolloin beeta-tyyppinen jaettu alatyyppeihin: beeta1-reseptorit (sydän, rasvasolujen) ja beeta-2-reseptoreihin (keuhkoputket, verisuonet, jne ...). Toiminnan isoproterenolin on beeta-1-reseptorin erinomainen toiminta adrenaliini ja noradrenaliini vain 10 kertaa, kun taas beeta-2-reseptoreihin se toimii 100-1000 kertaa voimakkaampi kuin luonnon katekoliamiinien.
Sovelluskohtaiset antagonistit (fenoksibentsamiini ja fentolamiini vastaan alfa- ja propranololi vastaan beeta-reseptori) vahvisti riittävyys luokittelun adrenoseptoreiden. Dopamiini pystyy olemaan vuorovaikutuksessa sekä alfa- ja beeta-reseptorien, mutta eri kudoksissa (aivoissa, aivolisäke, alusta) havaittu ja oma dopaminergisiin reseptoreihin erityinen salpaaja, joka on haloperidoli. Beetaseptorien määrä vaihtelee välillä 1000-2000 solua kohden. Biologiset vaikutukset katekoliamiinien välittämien beeta-reseptorit ovat liittyneet, pääsääntöisesti, jossa adenylaattisyklaasin aktivoitumisen ja solunsisäinen cAMP. Reseptori ja entsyymi, vaikka ne ovat toiminnallisesti liitettyjä, mutta edustavat erilaisia makromolekyylejä. Modulointi adenylaattisyklaasiaktiivisuuden vaikutuksen alaisena hormonin reseptorin kompleksin ovat mukana guanosiinitrifosfaatin (GTP) ja muut puriininukleotidien. Lisäämällä entsyymin aktiivisuutta, ne näyttävät vähentävän affiniteettia beeta-reseptorin agonisteja.
Denervoitujen rakenteiden herkkyyden lisäämistä koskeva ilmiö on pitkään tunnettu. Sitä vastoin pitkäaikainen altistus agonisteille pienentää kohdekudosten herkkyyttä. Beetasäteilijöiden tutkimuksen avulla voidaan selittää nämä ilmiöt. Osoitettiin, että isoproterenolin pitkäaikainen vaikutus johtaa adenylaattisyklaasin herkkyyden heikkenemiseen, mikä johtuu beetareseptorien määrän vähenemisestä.
Desensitisointiprosessi ei vaadi proteiinisynteesin aktivoitumista, ja se johtuu todennäköisesti palautumattomien hormonireseptorikompleksien asteittaisesta muodostumisesta. Päinvastoin, 6-oksidofamidin antaminen, joka rikkoo sympaattisia päitä, liittyy siihen, että kudoksissa on reagoivien beetareseptorien määrä lisääntynyt. Ei ole poissuljettua, että sympaattisen hermoston aktiivisuuden lisääntyminen määrää verisuonien ja rasvakudoksen ikääntymisen desensitisoinnin suhteessa katekoliamiineihin.
Eri elinten adrenoreseptoreiden määrää voidaan hallita muilla hormoneilla. Siten, se lisää estradioli, progesteroni ja vähentää alfa-adrenergisiin reseptoreihin kohtuun, johon liittyy vastaava lisäys ja vähennys sen supistusvastetta katekoliamiinien. Jos solunsisäinen "toisiolähetti", joka on muodostettu toiminnan beeta-reseptorin agonisteja, on varmasti cAMP, suhteessa lähetin alfa-adrenergisen vaikutukset on monimutkaisempi. Olemassaolo erilaisten mekanismien: vähentäminen cAMP-tasojen, cAMP-tasojen lisääminen modulaatio kalsiumin solujen dynamiikka, ja muut.
Toistaa erilaisia vaikutuksia kehossa tarvitaan yleensä adrenaliiniannoksella, on 5-10 kertaa pienempi kuin noradrenaliinin. Vaikka jälkimmäinen on tehokkaampi suhteessa a- ja beeta-1-adrenergiset reseptorit, on tärkeää muistaa, että endogeenisiä sekä katekoliamiinien voivat olla vuorovaikutuksessa sekä alfa- ja beeta-reseptoreihin. Näin ollen, biologista vastetta kehon adrenergisiin aktivointi pitkälti riippuu reseptorin siinä läsnä. Kuitenkin, tämä ei tarkoita, että valikoiva aktivointi hermo tai humoraalisen sympaattinen-lisämunuaisen järjestelmä on mahdotonta. Useimmissa tapauksissa sen eri yhteyksien tehostettu toiminta. Näin ollen oletetaan, että se aktivoi refleksi hypoglykemia lisämunuaisen ydin, kun taas verenpaineen lasku (posturaalinen hypotensio) mukana pääasiassa noradrenaliinin vapautumista sympaattisen hermoston päätteitä.
Adrenoreseptorit ja niiden aktivaation vaikutukset erilaisiin kudoksiin
|
Järjestelmä, elin |
Adrenoseptorin tyyppi |
Reaktio |
|
Sydän- ja verisuonijärjestelmä: |
||
|
Sydän |
Beeta |
Kontraktioiden, johtumisen ja supistumisnopeuden lisääntyminen |
|
Arterioleja: |
||
|
Iho ja limakalvot |
Alfa |
Vähentäminen |
|
Luurankolihaksia |
Beeta |
Laajennuksen vähennys |
|
Vatsan elimiä |
Alfa (lisää) |
Vähentäminen |
|
Beeta |
Laajentaminen |
|
|
Wien |
Alfa |
Vähentäminen |
|
Hengityselimet: |
||
|
Keuhkoputkien lihakset |
Beeta |
Laajentaminen |
|
Ruoansulatusjärjestelmä: |
||
|
Vatsa |
Beeta |
Moottorin vähentynyt moottoritoiminto |
|
Suolet |
Alfa |
Sphinserien vähentäminen |
|
Perna |
Alfa |
Vähentäminen |
|
Beeta |
Rentoutuminen |
|
|
Ulkopuolisesti salamainen osa haimasta |
Alfa |
Vähentynyt eritys |
|
Genitourinary järjestelmä: |
Alfa |
Sfingerien vähentäminen |
|
Rakko |
Beeta |
Rentouttava eksouristinen lihas |
|
Mies sukupuolielimiä |
Alfa |
Siemensyöksy |
|
Katse |
Alfa |
Oppilas laajentui |
|
Nahka |
Alfa |
Lisääntynyt hikoilu |
|
Salivaryhmät |
Alfa |
Kaliumin ja veden eristäminen |
|
Beeta |
Amylaasin erittyminen |
|
|
Hormonaaliset rauhaset: |
||
|
Haiman saarekkeet |
||
|
Beetasolut |
Alfa (lisää) |
Vähentynyt insuliinin erittyminen |
|
Beeta |
Lisääntynyt insuliinin erittyminen |
|
|
Alfa-solut |
Beeta |
Glukagonin lisääntynyt erittyminen |
|
8-solu |
Beeta |
Somatostatiinin lisääntynyt eritys |
|
Hypotalamuksen ja aivolisäkkeen: |
||
|
Somatotrofы |
Alfa |
STH: n lisääntynyt erittyminen |
|
Beeta |
STH: n vähentynyt eritys |
|
|
Laktotrofы |
Alfa |
Prolaktiinin väheneminen |
|
Tireotrofy |
Alfa |
TSH: n vähentynyt erittyminen |
|
Kortikotrofy |
Alfa |
ACTH: n lisääntynyt erittyminen |
| beeta | ACTH: n vähentynyt erittyminen | |
|
Kilpirauhanen: |
||
|
Follikulaariset solut |
Alfa |
Tyroksiinin vähentynyt erittyminen |
|
Beeta |
Tyroksiinin lisääntynyt erittyminen |
|
|
Parafollikulaariset (K) -solut |
Beeta |
Lisääntynyt kalsitoniinin erittyminen |
|
Lisäkilpirauhaset |
Beeta |
PTH: n lisääntynyt erittyminen |
|
Munuaiset |
Beeta |
Lisääntynyt reniinin eritys |
|
Vatsa |
Beeta |
Lisää gastriinin eritystä |
|
Perusvaihto |
Beeta |
Lisääntynyt hapen kulutus |
|
Maksa |
? |
Glykogenolyysi- ja glukoneogeneesin lisääntyminen glukoosin saannosta; lisätä ketogeneesiä vapauttamalla ketoni-elimet |
|
Rasvakudos |
Beeta |
Lipolyysin lisääntyminen vapaiden rasvahappojen ja glyserolin vapautumisella |
|
Luuston lihakset |
Beeta |
Glykolyysin lisääntyminen pyruvaatin ja laktaatin vapautumisen kanssa; proteolyysin väheneminen alaniinin, glutamiinin, saannolla |
On tärkeää muistaa, että tulokset laskimoon katekoliamiinien eivät aina heijasta riittävästi vaikutuksia sisäsyntyisen aineen. Tämä koskee lähinnä noradrenaliinin, koska se on kehon useimmiten ei veri, vaan suoraan synapsiraossa. Sen vuoksi endogeenisen norepinefriinin aktivoi, esimerkiksi, ei vain verisuonten alfa-reseptoreihin (verenpaineen nousu), mutta myös sydämen beeta-reseptoreihin (sydämentykytys), kun taas anto noradrenaliinin ulkoisen johtaa pääasiassa aktivoitumista verisuonten alfa-reseptorin ja refleksi (kautta vagus) hidastuminen sydämenlyönnit.
Pienet epinefriini-annokset aktivoivat lähinnä lihasastioiden ja sydämen beeta-reseptorit, mikä johtaa perifeerisen verisuonten resistenssin laskuun ja lisää sydämen minuuttitilavuutta. Joissakin tapauksissa ensimmäinen vaikutus voi vallita, ja adrenaliinin antamisen jälkeen hypotensio kehittyy. Suuremmilla annoksilla adrenaliini ja aktivoi alfa-reseptoreihin liittyy lisääntynyt ääreisverenkierron vastusta ja sydämen tausta kasvu johtaa verenpaineen nousu. Kuitenkin sen vaikutus vaskulaarisiin beetareseptoreihin säilyy. Tämän seurauksena systolisen paineen nousu ylittää samanarvoisen diastolisen paineen (pulssipaineen nousu). Ottamalla käyttöön edes suurina annoksina alkavat dominoida alfa-mimeettisestä vaikutukset adrenaliinia: systolinen ja diastolinen verenpaine kasvoi samaan, vaikutuksen alaisena noradrenaliinin.
Katekoliamiinien vaikutus aineenvaihduntaan muodostuu niiden suorista ja epäsuorista vaikutuksista. Ensimmäiset toteutetaan pääasiassa beta-reseptoreilla. Maksa liittyy monimutkaisempaan prosessiin. Vaikka maksan glykogenolyysin tehostumista on perinteisesti pidetty beeta-reseptorin aktivaation tuloksena, on myös tietoja a-reseptorien osallistumisesta tähän. Katekoliamiinien välitetyt vaikutukset liittyvät monien muiden hormonien, esimerkiksi insuliinin, erityksen modulaatioon. Toiminnassa eritykseen adrenaliini on selvästi hallitseva alfa-adrenergisen aineen, kuten on esitetty, stressiä, joka liittyy estämällä insuliinin eritystä.
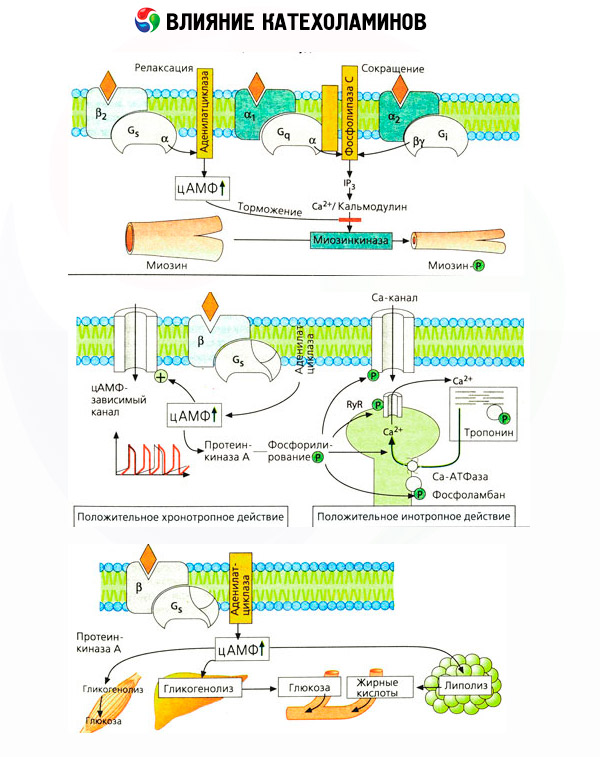
Yhdistelmä suorat ja epäsuorat vaikutukset katekoliamiinien johtaa hyperglykemiaan, konjugaatin paitsi kasvua maksan glukoosin tuotantoa, mutta myös inhibition kanssa sen käyttö Ääreiskudoksissa. Kiihtyvyys lipolyysin on giperlipatsidemiyu lisääntyneet toimitukset rasvahappojen maksaan, ja tehostamista tuotannon ketoaineiden. Vahvistaminen glykolyysin lihaksissa johtaa tuotanto kasvaa veren laktaattia ja pyruvaatti, joka yhdessä vapautunut glyseroli rasvakudoksesta, ovat edeltäjien maksan glukoneogeneesin.
Sääntely erityksen katekoliamiinien. Samankaltaisuus tuotteita ja menetelmiä vasteen sympaattisen hermoston ja lisämunuaisen ydin on perustana yhdistämällä nämä rakenteet yhdeksi elin sympatoadrenaalisen release neuraalisen ja hormonaalista sen linkki. Eri tuovissa signaalit ovat keskittyneet hypotalamuksen ja keskusten selkäytimen ja ydinjatkos välillä, jotka virtaavat vievissä paketti kytketään päälle preganglionic hermosolukuolemia elimet sijaitsevat sivusuunnassa sarvi selkäytimen tasolla kohdunkaulan VIII - II-III lannerangan segmenttejä.
Preganglionic aksonien näistä soluista lähtevät selkäytimen ja muodostavat synaptisten yhteyksien kanssa neuronien lokalisoitu ganglioiden sympaattisen ketjun, tai lisämunuaisytimessä soluja. Nämä ovat kolinergisiä preganglionic kuituja. Ensimmäinen perustavanlaatuinen ero sympaattiset ganglioiden jälkeiset neuronit ja lisämunuaisen ydin kromaffiinisoluissa koostuu siitä, että jälkimmäinen lähetetään saapuvan signaalin se kolinergisten neuro- johtuminen (postganglionic adrenerginen hermoja) ja humoraalisen korostamalla adrenergisen yhdisteen verta. Toinen ero pienenee postganglionic hermoja, jotka tuottavat noradrenaliinin, kun taas lisämunuaisytimessä solut - edullisesti adrenaliini. Nämä kaksi ainetta on erilaisia vaikutuksia kankaan.