
Aivolisäke
Viimeksi tarkistettu: 23.04.2024

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
Aivolisäkkeen (aivolisäkkeen, s.glandula pituitaria) tallennetaan aivolisäkkeen fossa sella seulaluussa ja kallon sisään, erotetaan kiinteä aine lisäke aivot kalvo, joka muodostaa kalvon istuin. Tämän kalvon reiän kautta aivolisäke on yhdistetty keskivartalon hypotalamuksen suppiloon. Aivolisäkkeen poikittainen koko on 10-17 mm, anteroposterior - 5-15 mm, pystysuora - 5-10 mm. Aivolisäkkeen paino miehillä on noin 0,5 g, naisilla 0,6 g. Ulkopuolella aivolisäke peitetään kapselilla.
Mukaisesti kehittämisessä aivolisäkkeen kahdesta eri bakteereita elimistössä erottaa kaksi osaa - edessä ja takana. Adenohypophysis tai etulohkon (adenohypophysis, s.lobus anterior), sitä suurempi on 70-80% kokonaispainosta aivolisäkkeen. Se on tiheämpi kuin takana oleva lohko. Kun etulohkon distaalisen osan eristetään (pars distalis), joka sijaitsee etuosan aivolisäkkeen fossa, väliosa (pars intermedia), joka on sijoitettu rajan takana osuus, ja bugornuyu osan (pars tuberalis), jolloin ylös ja liitetty suppiloon hypotalamuksessa. Johtuen runsaasti verisuonia etulohkon on vaalean keltainen väri ja punertava värisävy. Parenkyymi aivolisäkkeen etulohkon rauhas-, jota edustaa useita erilaisia soluja, jotka sijaitsevat nauhojen välillä sinimuotoinen kapillaareja. Puoli (50%) ja adenohypofyysissa solut ovat hromafilnymi adenocytes ottaa niiden sytoplasmassa hienoja rakeita, hyvin värjättiin kromin suoloja. Tämän acidophilic adenocytes (40% adenohypofyysissa solut), ja basofiilisiä adenocytes {10%). Määrä basofiilisia adenocytes kuuluvat gonadotrooppisina, kortikotropnye tyreotropiinille endocrinocytes. Kromofobiadenooma adenocytes pieniä, ne ovat suuri ydin ja pieni määrä sytoplasmaan. Näitä soluja pidetään kromofiilisten adenosyyttien prekursoreina. Muut 50% adenohypofiasolut ovat kromofobisia adenosyyttejä.
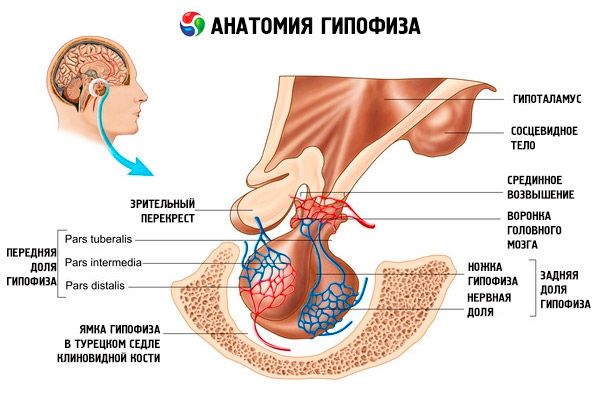
Neurohypophysis tai taka-lohko (neurohypophysis, s.lobus posteriorinen), joka koostuu hermo osa (lõbus nervosus), joka sijaitsee takaosassa aivolisäkkeen fossa ja suppilo (infundibulum) takana sijaitseva bugornoy osan adenohypophysis. Aivolisäkkeen taka-lohko muodostuu gliasoluista (aivolisäkkeen solut), hermo kuituja, jotka ulottuvat neurosekretorisen ytimet hypotalamus neurohypophysis ja neurosekretorisen soluja.
Aivolisäke hermokuitujen (reittien) ja verisuonten avulla on funktionaalisesti sidoksissa aivojen aivojen hypotalamukseen, joka säätelee aivolisäkkeen toimintaa. Aivolisäke ja hypotalamus yhdessä neuroendokriinisten, verisuoni- ja hermosolujensa kanssa pidetään yleensä hypotalamus-aivolisäkejärjestelmänä.
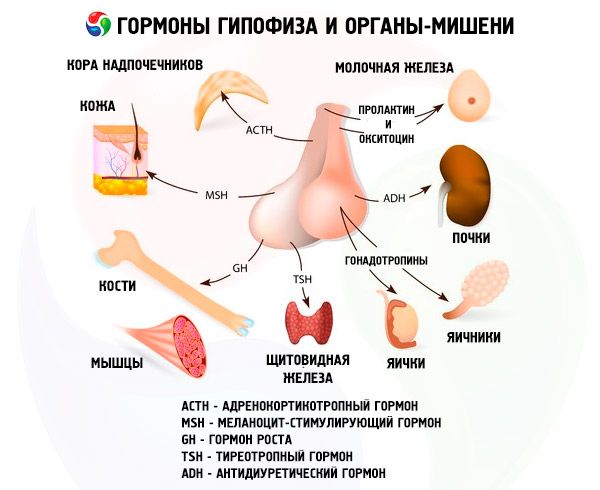
Hormonien etu- ja taka aivolisäkkeen vaikuttaa moniin kehon toimintoihin, etenkin muiden Umpieritysrauhaset. In etulohkon aivolisäkkeen acidophilic adenocytes (alfa) solut tuottavat somotropny hormonia (HGH), jotka osallistuvat säätelyssä kasvun ja kehityksen nuoren organismin. Kortikotropnye endocrinocytes erittävät adrenokortikotrooppisen hormonin (ACTH), joka stimuloi steroidihormonien lisämunuaiset. Tirotropnye endocrinocytes erittävät tirotropny hormoni (TSH), joka vaikuttaa kilpirauhanen kehitystä ja aktivoimalla tuotantoa sen hormoneja. Gonadotrooppisten hormonit: follikkelia stimuloiva hormoni (FSH), luteinisoivan hormonin (LH) ja prolaktiini - vaikuttaa murrosiän kehon, säännellä ja edistää kehitystä munarakkuloiden munasarjoissa, ovulaatio, rintojen kasvua ja maidontuotanto naisilla, prosessi spermatogeneesin miehillä. Näitä hormoneja tuottavat beta-solun basofiiliset adenosyytit ). Täällä lipotropic erittämät tekijät aivolisäkkeestä, mikä vaikuttaa mobilisointia ja hyödyntämiseen rasvaa kehossa. Väliosan etulohkon muodostettu melanosyyttejä stimuloiva hormoni, joka ohjaa pigmenttien muodostamisen - melaniinin - kehossa.
Hypotalamuksen supraoptisten ja paraventrikulaaristen ytimien neurosekulaariset solut tuottavat vasopressiiniä ja oksitosiinia. Nämä hormonit kuljetetaan aivolisäkkeen posteriorisen lohkon soluihin pitkin akseleja, jotka muodostavat hypotalamus-aivolisäkkeen. Aivolisäkkeen takajalusta nämä aineet tulevat veren sisään. Vasopressiinihormonilla on vasokonstriktori ja antidiureettinen vaikutus, jota kutsutaan myös antidiureettiseksi hormoniksi (ADH). Oksitosiini on stimuloiva vaikutus supistuvuuden lihakset kohtu, lisää maitoa imettävien maitorauhasen, inhiboi ja keltarauhastoimintaa, vaikuttaa tasainen muutos sävy (neischerchennyh) lihakset maha-suolikanavan.
Aivolisäkkeen kehittyminen
Aivolisäkkeen etuosa kehittyy oraalisen lahden takasolun epiteelistä rengasmaisen ulkonemisen muodossa (Rathke's tasku). Tämä ektodermaalinen ulkonema kasvaa tulevan III kammion pohjalle. Hänen kohti toisen aivorungon alapäästä (kolmannen kammion tulevaa pohjaa) kasvaa ulospäin, josta suppilon harmaa kraaterti ja aivolisäkkeen takajalusta kehittyvät.
 [1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
Aivolisäkkeen alukset ja hermot
Aivolisäkkeen ylempi ja alempi ateroskleroosi suuntautuvat aivolisäkkeen aivoverenkierron sisäisiltä kaulavaltimovirroilta ja verisuoneilta. Ylä hypophyseal valtimo menee harmaaksi tumaan suppiloon ja hypotalamuksen anastomoituvat keskenään täällä ja muodostavat tunkeutua aivokudokseen kapillaareja - gemokapillyarnuyu runkoverkon. Tämän verkon pitkien ja lyhyiden silmukoiden muodostami- sesta muodostuu portaalisuonet, jotka on suunnattu aivolisäkkeen etupäähän. Aivolisäkkeen etukannen parenkyymissä nämä suonet hajoavat laajaan sinimuotoiseen kapillaariin, jotka muodostavat toissijaisen hemokapillaryn verkon. Aivolisäkkeen takalohko on ensisijaisesti verenvuoto alemman aivolisäkkeen valtimon kautta. Aivolisäkkeiden ylä- ja alaosien välissä on pitkät valtimotukot. Laskimoverin ulosvirtaus toissijaisesta hemokapillaarisesta verkosta suoritetaan suonien järjestelmällä, joka virtaa aivojen kovan kuoren kavereisiin ja interdigitaalisiin sinusoireihin.
Aivolisäkkeen innervaatioon liittyy sympaattisia kuituja, jotka tunkeutuvat elimeen yhdessä verisuonien kanssa. Postganglionic sympaattiset hermokuidut siirtyvät pois sisäisen kaulavaltimon sisäpinnasta. Lisäksi aivolisäkkeen takareikään on löydetty lukuisia hypotalamuksen ytimissä sijaitsevien neurosekretoristen solujen proses- seja.
Aivolisäkkeen ikäominaisuudet
Aivolisäkkeen keskimääräinen paino vastasyntyneillä saavuttaa 0,12 g. Ruumiinpaino kaksinkertaistuu 10: een ja kolminkertaistuu 15 vuodella. Aivolisäkkeen paino saavuttaa enintään 20-vuotiaana (530-560 mg) ja myöhemmissä ikäkausina se lähes muuttuu. 60 vuoden kuluttua tämän hormonihoidon massa on vähentynyt hieman.
[14], [15], [16], [17], [18], [19], [20], [21], [22], [23], [24], [25], [26]
Aivolisäkkeen hormonit
Hermo- ja hormonaalisen säätelyn yhtenäisyys kehossa varmistetaan aivolisäkkeen ja hypotalamuksen läheisellä anatomisella ja toiminnallisella liitoksella. Tämä kompleksi määrittää koko endokriinisen järjestelmän tilan ja toiminnan.
Tärkeimmät umpirauhanen, joka tuottaa useita peptidihormonit suoraan säädellä toimintaa reuna rauhasten - aivolisäkkeestä. Se on punertavan harmaa papu-muotoinen muodostumista, peitetty sidekudoskotelosta jotka painoivat 0,5-0,6 g se vaihtelee hieman riippuen sukupuolen ja iän henkilö. On yleisesti hyväksyttyä, jako aivolisäkkeen kahteen osaan, erilaisia kehitys, rakenne ja toiminta: edessä distaalinen - anteriorisen aivolisäkkeen ja takaisin - neurohypophysis. Ensimmäinen noin 70% kokonaispainosta eturauhasen ja on jaettu distaalinen, Voronkov ja väliosa, toinen - on takaosa, tai osa, ja aivolisäkkeen varsi. Rauhanen sijaitsee aivolisäkkeen fossa sella seulaluussa ja haaran kautta on kytketty aivoihin. Etusuoran yläosa peitetään visuaalisella risteyksellä ja visuaalisilla reiteillä. Perfuusio aivolisäkkeen erittäin runsas oksat ja suoritetaan sisäisen kaulavaltimon (ylempi ja alempi aivolisäkkeen valtimoissa), ja oksat aivovaltimon ympyrä. Ylempi Aivolisäkkeen valtimon verenkierron mukana adenohypophysis ja alemman - neurohypophysis, jossa se saatetaan kosketukseen neurosekretorisen Axon päätteitä hypotalamuksen magnosellulaarisessa. Ensimmäinen osa mediaani paremmuus hypotalamuksen, jotka ovat levittäytyneet kapillaariverkon (ensisijainen Hiussuonipunoksen). Näiden kapillaarien (joka koskettimia aksonien pieni mediobasal hypotalamuksen neurosekretorinen solut) kerättiin porttilaskimon laskeva pitkin jalat aivolisäkkeen adenohypophysis peruskudoksen, jossa jälleen jaettu sinimuotoinen hiussuonia ketju (toissijainen Hiussuonipunoksen). Siten, veri, jotka on aikaisemmin viety mediaani hypotalamuksen eminentia jossa adenogipofizotropnymi rikastettu hypotalamuksen hormonia (kortikotropiinia vapauttava hormoni), saa adenohypofyysissa.
Ulosvirtaus veren, tyydyttynyt adenogipofizarnymi hormonit lukuisia kapillaareja toissijainen plexus laskimot suoritetaan järjestelmän, joka puolestaan virtaavat laskimoiden poskionteloiden ja kovakalvon ja verenkiertoon. Siten, aivolisäkkeen portaalin järjestelmän laskeva suunta veren virtauksen hypotalamuksen on osa monimutkaista mekanismia morphofunctional Neurohumoraalisten tropic ohjaustoiminnot adenohypophysis.
Aivolisäkkeen innervaatio suoritetaan sympaattisilla kuiduilla, jotka seuraavat aivolisäkkeen valtimoita. Alusta alkaen heille annetaan postganglionisia kuituja, jotka kulkevat sisäisen karotidiplexuksen kautta, joka liittyy ylempiin kohdunkaulan solmukohtiin. Adenohypöfyysistä ei ole suoraa innervaatiota hypotalamuksesta. Hypotalamuksen neurosekretoristen ytimien hermovärit tulevat takajalkoon.
Adenohypofysiikka histologisessa arkkitehtuurissa on hyvin monimutkainen muodostuminen. Se erottaa kahden tyyppiset rauhasten solut - kromofobiset ja kromofiiliset. Nämä puolestaan jaetaan eosinofiilien ja basofiilien (aivolisäkkeen yksityiskohtainen histologinen kuvaus annetaan sopiva käsikirjan osaa). On kuitenkin syytä huomata, että hormonit rauhasten soluja, jotka muodostavat peruskudoksen adenohypophysis, koska viime monimuotoisuuden jonkin verran erilaisia niiden kemiallisesta luonteesta ja hieno rakenne sekretiziruyuschih solujen olisi vastattava biosynteesin ominaisuuksia kunkin. Mutta joskus voi katsella adenohypophysis ja välimuotoja rauhas soluja, jotka pystyvät tuottamaan enemmän hormoneja. On näyttöä siitä, että erilaisia adenohypofysiikan rauhasten soluja ei aina määritellä geneettisesti.
Turkin satulan kalvon alapuolella on etusuoran suppilo-osa. Se kattaa aivolisäkkeen jalat ja koskettaa harmaata kukkulaa. Tämä osa adenohypophysis on luonteenomaista sen läsnäolo epiteelisolujen ja runsaasti verenkiertoa. Se on myös hormoniaktiivinen.
Aivolisäkkeen välivaihe (keskiosa) koostuu useista kerroksista suuria eritystä omaavia basofiilisiä soluja.
Aivolisäke hormonien läpi kuljettaa erilaisia toimintoja. Sen etulohkon tuotettu adrenocorticotropin (ACTH), kilpirauhasta stimuloiva (TSH), follikkelia stimuloiva hormoni (FSH), luteinisoiva hormoni (LH), lipotropic hormoni ja kasvuhormonin -. Somatotropic (SRT ja prolaktiini välituotteiden lohko syntetisoidaan melanosyyttejä stimuloiva hormoni (MSH), ja Takapuolessa kerääntyy vasopressiini ja oksitosiini.
AKTG
Hyphyseaaliset hormonit edustavat proteiinin ja peptidihormonien ryhmää ja glykoproteiineja. Aivolisäkkeen eturaajojen hormoneista ACTH on eniten tutkittu. Sitä tuottaa basophil-solut. Sen tärkein fysiologinen tehtävä on biosynteesin stimulointi ja steroidihormonien erittyminen lisämunuaiskuoressa. ACTH: llä on myös melanosyyttejä stimuloiva ja lipotrooppinen aktiivisuus. Vuonna 1953 se eristettiin puhtaassa muodossaan. Myöhemmin sen kemiallinen rakenne perustettiin, joka koostui 39 aminohappotähteestä ihmisessä ja useissa nisäkkäissä. ACTH: llä ei ole erityistä spesifisyyttä. Tällä hetkellä kemiallisen synteesin sekä itse hormoni että erilaiset, aktiivisempi kuin luonnolliset hormonit, sen molekyylin fragmentit toteutetaan. Hormonin rakenteessa peptidiketjun kaksi osaa, joista toinen antaa ACTH: n havaitsemisen ja sitoutumisen reseptoriin, ja toinen - antaa biologisen vaikutuksen. ACTH-reseptorilla vaikuttaa siltä, että se sitoutuu hormonin ja reseptorin sähköisten latausten vuorovaikutuksesta. Biologisen efektorin rooli ACTH suorittaa molekyylin 4-10 (Met-Glu-Gis-Fen-Arg-Tri-Tri) fragmentin.
ACTH: n melanosyyttistä stimuloiva aktiivisuus johtuu N-terminaalisen alueen molekyylistä, joka koostuu 13 aminohappotähteestä ja toistaa alfa-melanosyytti-stimuloivan hormonin rakennetta. Sama sivusto sisältää heptapeptidiä, joka on läsnä muissa aivolisäkkeissä ja jolla on joitain adrenokortikotrooppisia, melanoosyttistä stimuloivia ja lipotrooppisia vaikutuksia.
Keskeinen kohta toiminnan ACTH pidettävä aktivointi entsyymin proteiinin sytoplasmassa cAMP. Fosforyloitu proteiinikinaasi aktivoi entsyymiesteraasi, joka muuntaa kolesteroliesterit vapaaksi aineeksi rasvaisissa pisaroissa. Proteiini syntetisoidaan sytoplasmassa seurauksena fosforylaation ribosomaalisen vapaan kolesterolin stimuloi sytokromi P-450 ja siirtää sen lipidipisaroista mitokondrioissa, jossa on kaikki entsyymit varmistamiseksi kolesterolin muuttaminen kortikosteroideja.
[27], [28], [29], [30], [31], [32], [33], [34], [35], [36], [37], [38], [39]
Thyrotropic hormoni
TSH - tirotropiini - kilpirauhasen kehityksen ja toiminnan pääasiallinen säätelijä, kilpirauhashormonien synteesi ja erittyminen. Tämä monimutkainen proteiini - glykoproteiini koostuu alfa - ja beta - alayksiköistä. Ensimmäisen alayksikön rakenne on sama kuin luteinisoivan hormonin alfa-alayksikkö. Lisäksi se sopii suurelta osin eri eläinlajeihin. Ihmisen TSH: n ihmisen beeta-alayksikössä olevien aminohappotähteiden sekvenssi decipheroidaan ja koostuu 119 aminohappotähteestä. Voidaan todeta, että ihmisen TSH: n ja karjan beeta-alayksiköt ovat monessa suhteessa samankaltaisia. Beeta-alayksikössä määritetään glykoproteiinihormonien biologisen aktiivisuuden biologiset ominaisuudet ja luonne. Se myös varmistaa hormonien vuorovaikutuksen reseptoreihin eri kohde-elimissä. Kuitenkin beta-alayksikkö useimmissa eläimissä osoittaa spesifistä aktiivisuutta vasta sen jälkeen, kun se on liittynyt alfa-alayksikköön toimimalla eräänlaisena hormonin aktivaattorina. Jälkimmäinen, jolla on sama todennäköisyys, indusoi beeta-alayksikön ominaisuuksia määrittävän luteinisoivan, follikkelia stimuloivan ja tyrotonisen vaikutuksen. Löytyneen samankaltaisuuden avulla voidaan päätellä, että nämä hormonit ovat peräisin evoluutiosta yhdestä yleisestä esiasteesta, beeta-alayksikkö määrittää hormonien immunologiset ominaisuudet. Oletetaan, että alfa-alayksikkö suojaa beeta-alayksikköä proteolyyttisten entsyymien vaikutuksesta ja myös helpottaa sen kuljetusta aivolisäkkeestä perifeerisiin kohde-elimiin.
Gonadotrooppiset hormonit
Gonadotropiineja esiintyy kehossa LH: n ja FSH: n muodossa. Näiden hormonien funktionaalinen tarkoitus vähenee yleensä lisääntymisprosessien aikaansaamiseksi molemmissa sukupuissa olevissa yksilöissä. Ne, kuten TTG, ovat monimutkaisia proteiineja - glykoproteiineja. FSH aiheuttaa follikkelien kypsymistä naaraissa munasarjoissa ja stimuloi spermatogeneesiä miehillä. LH aiheuttaa naaraissa follikkelia puhkeamisen keltaisen ruumiin muodostumisen ja stimuloi estrogeenien ja progesteronin erittymistä. Miehillä tämä sama hormoni kiihdyttää interstitiaalisen kudoksen kehittymistä ja androgeenien erittymistä. Gonadotropiinien vaikutukset ovat riippuvaisia toisistaan ja edistyvät synkronisesti.
Gonadotropiinin erityksen dynamiikka naisilla vaihtelee kuukautiskierron aikana ja sitä tutkitaan riittävän yksityiskohtaisesti. Syklin preovulatiivisessa (follikulaarisessa) faasissa LH: n sisältö on melko alhaisella tasolla ja FSH lisääntyy. Follikkelin kypsymisen eritys estradiolin kasvoi, mikä lisää tuotannon gonadotropiinien aivolisäkkeen ja ulkonäkö syklit sekä LH: n ja FSH: n jne. E., Sex steroidit stimuloivat gonadotropiinien erityksen.
Tällä hetkellä LH: n rakenne määritetään. Kuten TTG, se koostuu 2 alayksiköstä: a ja p. LH: n alfa-alayksikön rakenne eri eläinlajeissa on pääosin samansuuntainen, se vastaa TSH: n alfa-alayksikön rakennetta.
Rakenteen beeta-alayksikköä LH on selvästi erilainen rakenne TSH beeta-alayksikön, vaikka se on neljä yhtä osan peptidin ketju koostuu 4-5 aminohappotähdettä. TTG ne ovat paikallisia asemissa 27-31, 51-54, 65-68 ja 78-83. Koska beeta-alayksikön LH: n ja TSH määrittää spesifinen biologinen aktiivisuus hormoneja, voidaan olettaa, että homologiset alueet rakenteen LH: n ja TSH pitäisi antaa beeta-alayksikön alfa-alayksikön ja rakenteeltaan erilaisia tontteja - vastaa spesifisyyden biologisen aktiivisuuden hormoni.
Natiivi LH on erittäin vakaa proteolyyttisten entsyymien, mutta beeta-alayksikön on lohkeavat nopeasti kymotrypsiinin, ja kova-alayksikköä hydrolysoidaan entsyymin, ts. E. Se on suojaava tehtävä, joka estää pääsyn kymotrypsiinin peptidisidoksia.
Mitä tulee FSH: n kemialliseen rakenteeseen, tällä hetkellä tutkijat eivät ole saaneet lopullisia tuloksia. Aivan kuten LH, FSH koostuu kahdesta alayksiköstä, mutta FSH: n beeta-alayksikkö eroaa LH: n beeta-alayksiköstä.
Prolaktiini
Toistokehityksessä aktiivisesti osallistuu toinen hormoni, prolaktiini (laktogeeninen hormoni). Prolaktiinin tärkeimmät fysiologiset ominaisuudet nisäkkäissä ilmenevät nisäkäsrauhasten ja imetyksen kehittymisen, talirauhasten ja sisäelinten kasvun muodossa. Se edistää steroidien vaikutusta toissijaisiin seksuaalisiin ominaisuuksiin miehillä, stimuloi keltaisen ruumiin eritystä aktiivisuutta hiirillä ja rotilla ja osallistuu rasvan aineenvaihdunnan säätelyyn. Viime vuosina prolaktiinille kiinnitetään paljon huomiota äidin käyttäytymisen sääntelijänä, ja tämä polyfunktionaalisuus selittyy sen evoluutiokehityksellä. Se on yksi muinaisista aivolisäkkeistä ja se löytyy myös sammakkoeläimistä. Tällä hetkellä eräiden nisäkäslajien prolaktiinin rakenne on täysin purettu. Kuitenkin vasta äskettäin tiedemiehet ovat ilmaisseet epäilynsä tällaisen hormonin olemassaolosta ihmisillä. Monet uskoivat, että sen toiminta tapahtuu kasvuhormonilla. Nyt meillä on vakuuttavia todisteita prolaktiinin läsnäolosta ihmisillä ja osittain decipheroitua sen rakenteesta. Prolaktiini-reseptorit sitoutuvat aktiivisesti kasvuhormoniin ja istukan laktogeeniin, mikä ilmaisee kolmen hormonin yksittäisen vaikutus- mekanismin.
Somatotropina
Vielä laajempi toiminta-aste kuin prolaktiinilla on kasvuhormoni - somatotropiini. Samoin kuin prolaktiinin, se tuottaa adenohypöfyysin acidofiiliset solut. STG stimuloi luuston kehittymistä, aktivoi proteiinin biosynteesiä, antaa rasvaa mobilisoivan vaikutuksen, edistää kehon koon kasvua. Lisäksi hän koordinoi valuuttaprosesseja.
Hormonin osallistuminen jälkimmäiseen vahvistaa sen seikka, että aivolisäkkeen erittyminen kasvaa voimakkaasti, esimerkiksi veren sokeripitoisuuden vähenemisellä.
Tämän ihmisen hormonin kemiallinen rakenne on nyt täysin perustettu - 191 aminohappotähdettä. Sen ensisijainen rakenne on samanlainen kuin korion somatomamotropiinin tai istukan laktogeenin rakenne. Nämä tiedot osoittavat näiden kahden hormonin merkittävän evoluutioisen läheisyyden, vaikka niillä on eroja biologisessa aktiivisuudessa.
On korostettava kyseessä olevan hormonin erityistä spesifisyyttä - esimerkiksi eläinperäinen STH on inaktiivinen ihmisillä. Tämä johtuu sekä ihmisen ja eläimen STH-reseptorien reaktiosta että itse hormonin rakenteesta. Tällä hetkellä on meneillään tutkimuksia aktiivisten alueiden tunnistamiseksi monimutkaisessa STH-rakenteessa, jolla on biologista aktiivisuutta. Tutkimme molekyylien yksittäisiä fragmentteja, joilla on muita ominaisuuksia. Esimerkiksi ihmisen STH: n pepsiinin hydrolyysin jälkeen eristettiin peptidi, joka koostui 14 aminohappotähteestä ja vastasi molekyylin 31-44 alueesta. Hänellä ei ollut kasvun vaikutusta, mutta lipotrooppinen aktiivisuus oli merkittävästi parempi kuin natiivihormoni. Ihmisen kasvuhormoni, toisin kuin eläimille samanlainen hormoni, on merkittävä laktogeeninen aktiivisuus.
Monissa adenohypophysis syntetisoitu sekä peptidi ja proteiini aineet, joilla on rasvaa liikkeelle paneva aktiivisuutta, ja trooppinen aivolisäkkeen hormonit - ACTH, kasvuhormoni, TSH ja muut - on lipotropic vaikutus. Viime vuosina erityisesti beta- ja y-lipotrooppiset hormonit (LPG) on erotettu toisistaan. Laajimmin tutkituista biologisia ominaisuuksia beeta-nestekaasua, joka, sen lisäksi lipotropic aktiivisuus on myös melanosyyttejä, kortikotropinstimuliruyuschee ja hypokalseemista vaikutus ja antaa vaikutelman insuliinia.
Tällä hetkellä dekoodataan lampaan nestekaasun (90 aminohappotähdettä), lipotrooppisten hormonien ja nautaeläinten primäärirakennetta. Tällä hormonilla on spesifinen spesifisyys, vaikka beeta-nestekaasun keskiosan rakenne eri lajeissa on sama. Se määrittää hormonin biologiset ominaisuudet. Yksi tämän alueen fragmentista löytyy alfa-MSH: n, beeta-MSH: n, ACTH: n ja beeta-nestekaasun rakenteesta. On ehdotettu, että nämä hormonit ovat peräisin samasta esiasteesta evoluution aikana. Y-nestekaasulla on heikompi lipotrooppinen aktiivisuus kuin beeta-nestekaasu.
Melanosyyttejä stimuloiva hormoni
Tämä hormoni syntetisoituu väli- koru on aivolisäkkeen, sen biologinen funktio stimuloiva ihon melaniinia biosynteesin, koko kasvaa ja määrä pigmentoituja melanosyyttien ihosoluissa sammakkoeläimet. Näitä MSH-ominaisuuksia käytetään hormonin biologisessa testauksessa. On olemassa kahdenlaisia hormoneja: alfa ja beta-MSH. Osoitetaan, että alfa-MSH: lla ei ole spesifistä spesifisyyttä ja sillä on sama kemiallinen rakenne kaikissa nisäkkäissä. Sen molekyyli on peptidiketju, joka koostuu 13 aminohappotähteestä. Beeta-MSH sen sijaan on spesifinen spesifisyys, ja sen rakenne eroaa eri eläimistä. Useimmilla nisäkkäillä β-MSH-molekyyli koostuu 18 aminohappotähteestä, ja vain ihmisillä se on pitkänomainen aminopäästä neljän aminohappotähteen suhteen. On huomattava, että alfa-MSH: lla on jonkin verran adrenokortikotrooppista aktiivisuutta ja sen vaikutus eläinten ja ihmisten käyttäytymiseen on nyt osoitettu.
Oksitosiini ja vasopressiinin
Posterior lohko aivolisäkkeen kerääntyä vasopressiinin ja oksitosiinin, jotka on hypotalamuksessa synteti-: vasopressiinin - in neuronien supraoptic ytimen, ja oksitosiinin - paraventrikulyatornogo. Sitten ne siirretään aivolisäkkeelle. On korostettava, että hypotalamuksessa syntetisoidaan ensin vasopressiinihormonin prekursori. Samanaikaisesti valmistetaan 1 ja 2-tyypin neurofitsiniproteiini. Ensimmäinen sitoo oksitosiinia ja toista - vasopressiinia. Nämä kompleksit kulkeutuvat neurosekretorinen rakeet sytoplasmassa pitkin Axon ja saavuttaa aivolisäkkeen jossa hermosyitä päättyvät verisuonen seinämän ja rae sisältö vereen. Vasopressiini ja oksitosiini ovat ensimmäiset aivolisäkkeen hormonit, joilla on täysin vahvistettu aminohapposekvenssi. Kemiallisessa rakenteessaan ne ovat nonapeptidejä yhden disulfidisillan kanssa.
Pidetään hormonit tuottaa erilaisia biologisia vaikutuksia: edistää veden kuljetus ja suolat kalvon läpi, on verenpainetta kohottava vaikutus, lisätä supistuminen sileän lihaksen kohdun synnytyksen aikana, lisätä eritystä maitorauhasista. On huomattava, että vasopressiinin antidiureettinen aktiivisuus on suurempi kuin oksitosiini, kun taas jälkimmäinen vaikuttaa voimakkaammin kohtuun ja maitorauhaseen. Tärkein säätelijä eritystä vasopressiinin on veden munuaistiehyeissä, se sitoutuu reseptoreihin solukalvon ja myöhempi aktivaatio entsyymin adenylaattisyklaasin niihin. Hormonin sitomiseksi reseptoriin ja biologiseen vaikutukseen molekyylin eri osat ovat vastuussa.
Aivolisäkkeen liittyvät hypotalamuksen kautta hermostoon, yhdistää kokonaisluvun toiminnallinen hormonitoimintaa mukana varmistamassa pysyvyyden sisäinen ympäristö (homeostaasin). Sisällä hormonitoimintaa säätelyyn perustuu periaatteeseen palautteen välillä etulohkon aivolisäkkeen ja zhelezami- "targets" (kilpirauhasen, lisämunuaisen kuorikerros, sukuelimiin). Kohdeohoren tuottaman hormonin ylimäärä estää ja sen puute stimuloi vastaavan trooppisen hormonin erittymistä ja vapautumista. Palautelijärjestelmään kuuluu hypotalamus. Se on siinä herkkä rauta-kohteiden hormoneille, reseptorivyöhykkeille. Sitoutumaan spesifisesti verenkierrossa hormoneja veressä ja muuttamalla reaktio konsentraatiosta riippuen hormonien, hypotalamuksen reseptorit välittävät sen vaikutus asiaa hypotalamuksen keskusten että koordinoi aivolisäkkeen etulohkon, hypotalamuksen vapauttavat hormonit adenogipofizotropnye. Siten hypotalamusta tulisi pitää neuro-endokriinisena aivoina.