
Ihmisen immuunikatovirus (HIV)
Viimeksi tarkistettu: 23.04.2024

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
Hankittu immunodeficiooireyhtymä eristettiin erityisenä sairaudena vuonna 1981 Yhdysvalloissa, kun useissa nuorissa vakavia sairauksia aiheuttivat mikro-organismit, jotka eivät ole patogeenisiä tai joilla on vähän patogeenisiä terveille ihmisille. Potilaiden immuunitutkimus paljasti lymfosyyttien lukumäärän jyrkkää vähenemistä yleensä ja erityisesti T-avustajia. Tätä sairautta kutsutaan nimellä AIDS (englantilainen hankittu immuunijärjestelmän oireyhtymä - hankittu immuunikatovirus tai aids). Infektiomenetelmä (seksuaalinen kosketus vertaan ja sen valmisteiden kautta) osoitti taudin tarttuvan luonteen .
Aiheuttava aine aids löydettiin 1983 itsenäisesti ranskalainen L. Montagnier, joka sanoi sen LAV lymphoadenopathy- Associated Virus), koska havaittiin potilaassa lymfadenopatia; ja amerikkalainen R. Gallo, joka kutsui viruksen HTLV-III: n (englanninkielen ihmisen T-lymfotrooppinen virus III): aiemmin hänet havaittiin lymfotrooppisista viruksista I ja II.
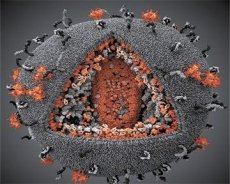
LAV- ja HTLV-III-virusten ominaisuuksien vertailu osoitti niiden identiteetin, joten vuonna 1986 virusta kutsuttiin HIV: ksi (ihmisen immuunikatovirus tai HIV) sekaannuksen välttämiseksi. HIV on pallomainen, sen halkaisija on 110 nm. Viruksen kirjekuoressa on polyhedron muoto, joka koostuu 12 pentagonista ja 20 kuusikulmasta. Glykosyloidun proteiinin gpl20 molekyyli sijaitsee kunkin kuusikulman keskiosassa ja kulmissa (numero 120 tarkoittaa proteiinin molekyylipainoa kilodaltonissa). Virion pinnalle on sijoitettu yhteensä 72 gpl20 molekyyliä, joista kukin liittyy intramembraaniproteiiniin gp41. Nämä proteiinit yhdessä kaksois lipidikerroksen kanssa muodostavat virionin superkapsidin (membraanin).
Proteiinit gpl20 ja gp41 muodostuvat esiasteproteiinin Env. Soluproteaasileikkauksen seurauksena. Proteiini gp41 muodostaa selkärangan, joka sytoplasminen domeeni yhdistää matriisiproteiinilla p17MA välittömästi kalvon alapuolella. Molion p17, joka vuorovaikuttaa virion kypsymisen kanssa, muodostaa kuosen taustalla olevan icosahedronin.
Virionin keskiosassa p24-proteiini muodostaa kartiomainen kapsidi. Kapselin kaventunut osa, johon pb-proteiini osallistuu, liittyy virionin kirjekuoreen. Kapsidissa on kaksi identtistä viruksen genomisen RNA: n molekyyliä. Ne sidotaan niiden 5'-päissä nukleokapsidiproteiiniin p7NC. Tämä proteiini on mielenkiintoinen, koska siinä on kaksi aminohappotähdettä (motiivi), kysteiini-rikas ja histidiini-atomi ja joka sisältää Zn, - niitä kutsutaan "sinkkisormi", koska ne kaapata molekyyli genomisen RNA: n liitettäväksi virioneihin muodostettu. Kapsidi sisältää myös kolmea entsyymiä. Reversase (RT) tai pol-kompleksi sisältää käänteistranskriptaasi, RNA-ase H ja DNA: sta riippuvainen DNA-polymeraasi. Revutaasi on läsnä p66 / p51-heterodimeerinä. Proteaasi (PR) - pI, käynnistää ja realisoi virion kypsymisen prosessin. Integraatio (IN) - p31 tai endonukleaasi takaa proviraalisen DNA: n sisällyttämisen isäntäsolun genomiin. Kapsidi sisältää myös siemen-RNA-molekyylin (tRNAl "3).
Solussa oleva RNA-geeni muunnetaan käänteistranskriptaasilla DNA-genomiin (DNA-provirus), joka koostuu 9283 nukleotidiparista. Se on rajoitettu vasempaan ja oikeaan ns. Pitkää toistetta tai LTR (englanti pitkä terminaalinen toisto): S'-LTR - vasen ja Z'-LTR - oikealla. LTR sisältää 638 nukleotidiparia.
HIV-genomi koostuu 9: sta geenistä, joista osa on päällekkäisiä (sillä on useita lukukehyksiä) ja sillä on eksoniinirakenne. He ohjaavat 9 rakenteellisen ja 6 säätelyproteiinin synteesiä.
Viruksen genomin LTR-arvo on se, että ne sisältävät seuraavat sääntelyelementit, jotka ohjaavat sen toimintaa:
- transkription signaali (promoottorialue);
- signaali poly-A: n lisäämiseksi;
- Kaappaussignaali;
- signaalin integrointi;
- positiivinen säätelysignaali (TAR TAT-proteiinille);
- negatiivisen sääntelyn elementti (NEF-proteiinin NRE);
- siemen-RNA: n (tRNA ™ 3) kiinnityspaikka DNA: n miinusketjun syntetisoimiseksi 3'-päässä; signaali LTR: n 5'-päähän, joka toimii alukkeena DNA: n plusviiran synteesille.
Lisäksi LTR sisältää elementtejä, jotka liittyvät mRNA: n liittämisen säätelyyn, pakkaamalla vRNA-molekyylit kapsidiin (Psi-elementti). Lopuksi, kun genomit kirjataan pitkäihin mRNA: iin, muodostetaan kaksi signaalia REV-proteiinille, joka vaihtaa proteiinisynteesiä: CAR säätelevien proteiinien ja CRS: n rakenteellisten proteiinien suhteen. Jos REV-proteiini sitoutuu CAR: hen, rakenteelliset proteiinit syntetisoidaan; jos se puuttuu, syntetisoidaan vain sääntelyproteiineja.
Viruksen genomin säätelemisessä on erityisen tärkeä rooli seuraavilla säätelevillä geeneillä ja niiden proteiineilla:
- TAT-proteiini, joka suorittaa positiivisen kontrollin viruksen lisääntymisen suhteen ja toimii säätelyalueella TAR;
- proteiineja NEV ja VPU, suorittaen negatiivisen kontrollin lisääntymisen NRE-kohdan kautta;
- proteiini REV, suorittaen positiivisen negatiivisen kontrollin. Protein REV ohjaa gag-, pol- ja env-geenien toimintaa ja tekee negatiivisen säätelyn yhdistämisen.
Näin ollen HIV: n lisääntyminen on kolmoisohjauksella - positiivinen, negatiivinen ja positiivinen-negatiivinen.
VIF-proteiini määrittää uuden syntetisoidun viruksen infektiivisyyden. Se sitoutuu kapsidiproteiiniin p24, ja se on läsnä virion määrässä 60 molekyyliä. NEF-proteiini on virion tapauksessa esitetty pienellä määrällä molekyylejä (5-10), mahdollisesti liitettynä kirjekuoreen.
VPR-proteiinia estää solusyklin G2-vaiheessa, kuljetuksessa preintegratsionnyh komplekseja solun tumassa ja aktivoi tiettyjen virusten ja solujen geenien, tehostaa viruksen replikaatiota monosyyteissä ja makrofageissa. Proteiinien VPR, TAT, REV, VPU virion sijaintia ei ole osoitettu.
Omien proteiinien lisäksi virion kalvon koostumus voi sisältää isäntäsolun joitakin proteiineja. Proteiinit VPU ja VPR osallistuvat viruksen lisääntymisen säätelyyn.
Ihmisen immuunikatoviruksen (HIV) antigeeniset variantit
Ihmisen immuunikatovirus (HIV) on hyvin vaihteleva. Jopa yhden potilaan organismeista voidaan erottaa viruksen kantoja, jotka eroavat merkittävästi antigeenisissä ominaisuuksissa. Tällaista vaihtelevuutta edistää CD4 + -solujen voimakas tuhoaminen ja voimakas vasta-ainevaste HIV-infektiolle. Länsi-Afrikan potilailla on uusi HIV-muoto, biologisesti lähellä HIV-1: tä, mutta immunologisesti erilainen HIV-2. Näiden virusten genomien primaarisen rakenteen homologia on 42%. DNA-provirus HIV-2 sisältää 9671 bp, ja sen LTR-854 bp. HIV-2 eristettiin tämän jälkeen muualla maailmassa. HIV-1: n ja HIV-2: n välillä ei ole ristiintuntemusta. Tunnetaan kaksi suurta HIV-1-muotoa: O (Outlier) ja M (Major), jälkimmäiset on jaettu 10 alatyyppiin (AJ). Venäjällä 8 alityyppiä liikkuu (AH).
 [
[HI-viruksen ja solun vuorovaikutusmekanismi
Kun tunkeutuu kehoon, virus tunkeutuu ensin soluihin, jotka sisältävät spesifisen CD4-reseptorin. Tällä reseptorilla on suuri määrä T-apuaineita, vähemmän makrofageissa ja monosyytteissä, erityisesti viruskeskeissä T-apuaineissa.
Ihmisen immuunikatovirus (HIV) tunnistaa CD4-reseptorit gpl20-proteiinillaan. Hivin ja solun vuorovaikutusprosessi etenee seuraavan kaavan mukaan: reseptorivälitteinen adsorptio -> rajautunut fovea -> rajautunut vesikkeli -> lysosomi. Siinä virionmembraani sulautuu lysosomikalvon kanssa, ja superkapsidista vapautunut nukleokapsidi tulee sytoplasmaan; matkalla tielle, joka tuhoutuu, ja genominen RNA ja siihen liittyvät ydinkomponentit vapautetaan. Seuraavaksi käänteistranskriptaasi syntetisoi on virionin RNA-miinus-DNA-juosteen, jonka jälkeen ribonukleaasi H tuhoaa virionin RNA: n ja virus-DNA-polymeraasi syntetisoi DNA: ta plus-ketjun. DNA-proviruksen päähän muodostuu 5'-LTR ja 3'-LTR. DNA-provirus voi olla ytimessä jonkin aikaa inaktiivisessa muodossa, mutta ennemmin tai myöhemmin se integroidaan integraasin avulla kohdesolun kromosomiin. Siinä provirus on inaktiivinen, kunnes tämä T-lymfosyytti aktivoidaan mikrobien antigeeneillä tai muilla immunokompetenteilla soluilla. Solu-DNA: n transkriptiota aktivoidaan säädellään erityisellä ydintekijällä (NF-kB). Se on DNA: ta sitova proteiini ja sitä tuotetaan suurina määrinä T-lymfosyyttien ja monosyyttien aktivaation ja proliferaation aikana. Tämä proteiini sitoutuu spesifisten solujen DNA-sekvenssien ja vastaavien LTR-DNA-provirus-sekvenssien sekvenssiin ja indusoi sekä solun DNA: n että DNA-proviruksen transkriptiota. DNA-proviruksen transkription indusoimalla hän siirtää viruksen inaktiivisesta tilasta aktiiviseksi ja siten jatkuvaksi infektioksi tuottavaksi. Pysy proviruksen ollessa passiivisessa tilassa voi kestää hyvin kauan. Viruksen aktivointi on kriittinen hetki vuorovaikutuksessa solun kanssa.
Siitä hetkestä lähtien, kun virus saapuu soluun, alkaa HIV-infektion aika, joka voi kestää 10 vuotta tai kauemmin; ja koska viruksen aktivaatio alkaa sairaudesta - aidsista. Heidän sääntelygeeniensa ja niiden tuotteiden avulla virusta alkaa moninkertaistaa aktiivisesti. TAT-proteiini voi lisätä viruksen lisääntymisnopeutta 1000 kertaa. Viruksen transkriptio on monimutkainen. Se sisältää sekä täyspitkän ja algenomisen mRNA: n muodostamisen, mRNA: n silmukoinnin ja rakenteellisten ja säätelevien proteiinien synteesin.
Rakenteellisten proteiinien synteesi tapahtuu seuraavasti. Ensiksi syntetisoidaan polyproteiinin edeltäjä Pr55Gag (proteiini, jonka massa on 55 kD). Se sisältää neljä päädomeenia: matriisi (MA), kapsidi- (CA), nukleokapsidi (NC) ja verkkotunnuksen pe, joka johtaa leikkaus Pr55Gag viruksen proteaasin (se samovyrezaetsya kohteesta toiseen proteiiniin esiaste - Gag-Pol) on muodostettu vastaavasti rakenteellisia proteiineja p17 , p24, p7 ja pb. Polyproteiinin Pr55Gag muodostaminen on tärkein edellytys viruspartikkeleiden muodostumiselle. Se on tämä proteiini, joka määrittää virion morfogeneesi-ohjelman. Se sisältää vaiheet, joissa peräkkäin Gag-polyproteiinin kuljetuksen plasmamembraanin ja sen vuorovaikutus proteiini-proteiini-vuorovaikutusten muodostumista viruspartikkelin ja sen orastava. Pr55Gag syntetisoidaan vapailla polyribosomeilla; proteiinimolekyylit kuljetetaan kalvoon, johon ne ankkuroituvat niiden hydrofobisiin laastareihin. Keskeinen rooli Gag-proteiinin luontaisen konformaation luomisessa CA-toimialueella. NC-domeeni kytkin tarjoaa (avulla "sinkkisormia") 2-molekyylin genomisen RNA viruspartikkelin muodostamiseksi. Polyproteiinimolekyyli ensin dimeroituu matriisidomeenien vuorovaikutuksen vuoksi. Sitten dimeerit yhdistetty heksameerinen (6 kpl) kompleksien vuorovaikutuksen domainit CA ja NC. Lopuksi, heksameerejä, yhdistävät sivupinnat muodostavat epäkypsä virionien pallomainen muoto, jonka sisällä sisältävät viruksen genomisesta RNA: sta jää NC-domeenin.
Toinen prekursoriproteiinia Prl60Gag-Pol (proteiini m. M. 160 kDa), syntetisoidaan seurauksena siirtää lukukehyksen ribosomin translaation aikana Z'-päähän gag-geenin alue välittömästi ennen koodaavan alueen RB-proteiinia. Tämän Gag-Pol-polyproteiinin käsittää osittaisen sekvenssin Gag-proteiinin (1-423 aminohappoa) ja Pol sekvenssit, jotka sisältävät domeeneja PR, RT: n ja IN. Polyproteiinin Gag-Pol molekyylejä syntetisoidaan myös vapailla polyribosomeilla ja kuljetetaan plasmamembraaniksi. Prl60Gagpol polyproteiinin sisältää kaikki tyypilliset sivustot polyproteiinin Gag välisten vuorovaikutusten ja sitoutumiskohtien kalvon. Näin ollen molekyylit polyproteiinin Gag-Pol sulake kalvon kanssa ja yhdessä Gag-molekyylejä ovat muodostavan virionit, joka voi johtaa aktiivisen proteaasin ja virionin kypsymisen prosessi alkaa. Proteaasin HIV-1 Erittäin aktiivinen vain dimeerinä, joten se samovyrezaniya on Prl60Gag-Pol vaatii dimerisaation näiden molekyylien. Virion kypsyminen on, että vapautunut aktiivinen proteaasi leikkaa prl60Gag-Pol: n ja Gag55: n tunnistettaviin kohteisiin; proteiinit p17, p24, p7, p6, revertase, integraasi muodostuvat ja ne assosioituvat virusrakenteeseen.
Env-proteiini syntetisoidaan liittyvät ribosomit limakalvojen Endoplasmakalvosto, glykosyloituja ja sitten se leikataan solun proteaasi gp120: een ja gp41 ja kuljetetaan solun pintaan. Tässä tapauksessa gp41 läpäisee membraanin ja sitoutuu Gag-proteiinimolekyylin matriisidomeeneihin, jotka liittyvät membraanin sisäpintaan. Tämä suhde pysyy kypsässä virion suhteen.
Siten, viruspartikkelien kokoamisen on aggregaatiota prekursoriproteiinien ja niihin liittyvät RNA-molekyylien solukalvon isäntäsolun, kypsymättömien virionien ja niiden vapautumista silmukoitumalla solun pinnalla. Kun Analysis virioni ympäröi itsensä solukalvon, joka on rakennettu gp41 ja gp120-molekyylin. Aikana jakautumisen ja mahdollisesti sen jälkeen, kun vapautuminen virionin kypsymisen tapahtuu, joka suoritetaan käyttäen viruksen proteaasin proteolyyttisen leikkaus Pr55Gag prekursoriproteiinien ja Prl60Gag-Pol virus kypsä proteiinit ja niiden yhdistyksen erityisiä rakenteellisia komplekseja. Johtavassa asemassa morphogeneesiä viruksen näyttelee polyproteiinin esiastetta Pr55Gag, joka järjestää ja kokoaa epäkypsä virionin; Sen kypsymisen prosessi täydentää spesifinen virusproteaasi.
Immunogeenisuuden syyt
Yksi HIV-infektion immuunikatavuuden pääasiallisista syistä on T-avustajien kuolema. Se tapahtuu seuraavien tapahtumien vuoksi. Ensinnäkin virustartunnan saaneista T-helper-viruksista apoptoosin takia. Uskotaan, että potilailla, joilla on AIDS-virus replikointi, apoptoosin ja väheneminen T-auttajasolujen ovat yhteydessä toisiinsa. Toiseksi, T-killer-solut tunnistavat ja tuhoavat T-solut, jotka on infektoitu viruksella, tai sisältää adsorboituneen gpl20-molekyyliä, samoin kuin virus-tartunnan ja virus-tartunnan auttaja-T-soluja, jotka muodostavat symplasts (synsytian), joka on useita kymmeniä soluja (osa he kuolevat virusten lisääntymisen seurauksena). Häviämisen vuoksi suuri määrä T-auttajasolujen on vähentynyt kalvon reseptoreiden ilmentymisen B-lymfosyyttien IL-2, häiriintynyt synteesi eri interleukiinien (kasvutekijöiden ja B-lymfosyyttien erilaistumi- - IL-4, IL-5, IL-6 et ai.), minkä seurauksena T-killer-järjestelmän toimintaa rikotaan. Täydennys- ja makrofagijärjestelmien toiminnan suppressaatio tapahtuu. Virusinfektoiduissa makrofagit ja monosyytit haluavat kuolla, mutta ne eivät pysty poistamaan virusta kehosta. Lopuksi, johtuen rakenteellisen ja antigeeninen samankaltaisuuksia reseptorien gpl20 joitakin epiteelisoluja organismin (kuten trofoblasteissa reseptoriin välittämisessä HIV-tartunnan siirteen) syntetisoidaan antiretseptornyh vasta-aineita, joilla on laaja kirjo toimintaa. Tällaiset vasta-aineet voivat estää erilaisia solu- reseptoreita ja mutkistaa taudin kulkua autoimmuunisairauksilla. Hiv-tartunnan seurauksena on immuunijärjestelmän kaikkien tärkeimpien osien häviäminen. Tällaiset potilaat muuttuvat puolustuksetta monenlaisia mikro-organismeja vastaan. Tämä johtaa opportunististen infektioiden ja neoplastisten tautien kehittymiseen. HIV-infektiota sairastavilla potilailla on ollut vähintään kolme syöpätaudin riskiä: Kaposin sarkooma; karsinoomat (mukaan lukien ihosyöpä); B-solulymfooma, joka johtuu B-lymfosyyttien pahanlaatuisesta degeneraatiosta. HIV: llä ei kuitenkaan ole vain lymfosyyttiä, vaan myös neurotrooppista. Se tunkeutuu CNS-soluissa (astrosyytit) joko reseptorivälitteisen endosytoosin ja fagosytoosin astrosyyttien virus-tartunnan lymfoblasteja. Vuorovaikutuksessa viruksen kanssa astrosyytit myös muodostettu symplasts edistä taudinaiheuttajan leviämisen solujen välisen kanavia. Makrofageissa ja monosyyteissä, virus voi säilyä pitkään, joten ne toimivat varastona ja sen jälleenmyyjien kehossa, joka kykenee tunkeutumaan kaikkiin kudoksiin. Tartunnan saaneilla makrofageilla on suuri merkitys HIV: n kulkeutumisessa keskushermostoon ja sen tappioon. Kymmenessä prosentissa potilaista primaariset kliiniset oireet liittyvät CNS-vaurioon ja ilmenevät dementiana (dementia). Niinpä hiv-tartunnan saaneille ihmisille on 3 sairausryhmää - opportunistiset infektiot, kasvainsairaudet ja keskushermoston vauriot.
HIV-infektion epidemiologia
HIV-infektion lähde on vain henkilö - sairas tai virustorjunta. Ihmisen immuunikatovirus (HIV) esiintyy veressä, siemennesteessä, kohdunkaulanesteessä; imettäville äideille - äidinmaidossa. Infektio tapahtuu seksuaalisesti, veren ja sen lääkkeiden sekä äidin ja lapsen välillä ennen synnytystä, synnytyksen aikana ja sen jälkeen. Virustartunnan tapauksia ruoan, juomien ja hyönteisten puremien kautta ei tunneta.
Huumeriippuvuus edistää aidsin leviämistä. HIV-infektio kasvaa joka vuosi. WHO: n mukaan vuosina 1980-2000 HIV: stä tartunnan saaneita oli 58 miljoonaa. Vain vuonna 2000 maailmassa sai tartunnan saaneita 5,3 miljoonaa ihmistä, ja AIDSista kuoli 3 miljoonaa ihmistä. Venäjällä 1. Tammikuuta 2004 lähtien oli rekisteröitynyt 264 000 HIV-positiivista henkilöä. Puolet HIV-tartunnan saaneista kuolee 11-12 vuoden kuluessa infektiosta. Vuoden 2004 alkupuolella 100 000 venäläisestä kansalaisesta noin 180 ihmistä elivät diagnosoimalla "HIV-infektio". On ennustettu, että HIV-tartunnan saaneiden ihmisten kokonaismäärä Venäjällä vuoteen 2012 mennessä on 2,5-3 miljoonaa ihmistä. HIV-infektion torjunnan monimutkaisuus riippuu useista syistä: ensinnäkin sen hoitoon ja erityiseen ehkäisyyn ei ole tehokkaita menetelmiä; Toiseksi, HIV-infektion inkubaatioaika voi olla yli 10 vuotta. Sen kesto riippuu T-lymfosyytin ja sen kromosomin sisältämän DNA-proviruksen aktivaation hetkestä. Ei ole vielä selvää, onko jokainen AIDS: llä tartunnan saaneilla virus ole tuomittu tai todennäköisesti on pitkäkestoinen virus ilman tautia (joka näyttää epätodennäköiseltä). Lopuksi on olemassa useita ihmisen immuunikatoviruksia (HIV-1, HIV-2), joiden antigeeniset erot estävät ristikkäimmän immuniteetin muodostumisen. Apinoiden immuunikatoviruksen (SIV) havaitseminen paljastaa HIV: n alkuperän. Genomin järjestämisen SIO on samanlainen kuin HIV, mutta se eroaa merkittävästi nukleotidisekvenssissä. HIV-2 serologisesti miehittää välituotteen HIV-1: n ja SIV: n välillä ja nukleotidisekvenssi oli lähempänä SIV: ää. Tässä suhteessa VM Zhdanov ehdotti, että HIV-1, HIV-2 ja SIV-virukset ovat peräisin yhteisestä esi-isästä. On mahdollista, mukaan R. Gallo, yksi SIV jotenkin joutui ihmiskehoon, jolle on tehty useita mutaatioita, jotka johtavat mihin tahansa HIV-1, HIV-2 ja sen muita muotoja.
HIV-infektion oireet
Ihmisen immuunipuutos-virukseen on ominaista tiettyjä ominaisuuksia, joihin taudin patogeneesi riippuu suurelta osin. Virus on erittäin korkea lisääntymisaste, jonka määräytyvät sen säätelyelementit (5 000 virion syntetisoidaan 5 minuutin kuluessa aktiivisessa vaiheessa). Fuusioproteiinin (gp41) läsnäolon vuoksi virus indusoi laajan syncysirakenteiden muodostumista infektoituneiden ja ei-infektoituneiden T-helpottajien yhdistämisen vuoksi, mikä johtaa niiden massakuolemaan. Suuret molekyyliset gpl20-molekyylit vapaasti kiertävät veressä ja sitoutuvat infektoitumattomien T-avustajien reseptoreihin, minkä seurauksena T-tappajat tunnistavat ja tuhoavat ne. Virus voi levitä solujen välisten solujen välityksellä solusta soluun, jolloin tässä tapauksessa vasta-aineet ovat vähemmän saatavilla.
HIV-infektion kliiniset kriteerit
Aikuisten HIV perustaa, jos heillä on ainakin kaksi vakavaa oireita yhdessä vähintään yhden oireen Alaikäisen ilman muiden tunnettujen syiden immuunipuutos (syöpä, synnynnäinen immuunipuutos, vaikea nälkään, ja niin edelleen. P.). Vakavia oireita ovat:
- laihtuminen 10%: lla tai enemmän;
- pitkittynyt kuume, ajoittainen tai pysyvä;
- krooninen ripuli.
Lievät oireet ovat itsepintainen yskä, yleistynyt ihottuma, toistuva herpes zoster, kandidiaasi suuontelon ja nielun, krooninen herpes simplex, yleinen lymfadenopatia. Aidsin diagnoosi tehdään vain Kaposin sarkooman, kryptokokin aivokalvontulehduksen, pneumocystis-keuhkokuumeen läsnäollessa. Tautien kliinistä kuvaa vaikuttaa opportunistiseen infektioon.
Ihmisen immuunikatoviruksen (HIV) viljelymenetelmät
HIV-1: tä ja HIV-2: ta voidaan viljellä vain yhden TCB4-lymfosyyttien - H9 soluissa, jotka on saatu leukemiallisista TCV4-lymfosyyteistä. Astroyyttisolujen monolayer-viljelmiä voidaan myös käyttää näihin tarkoituksiin, joissa HIV-1 kertoo hyvin. Eläimistä hiv-1: hen herkkiä simpansseja.
Viruksen resistanssi ulkoisessa ympäristössä on vähäinen. Hän kuolee auringonvalon ja UV-säteilytyksen vaikutuksesta, tuhotaan 80 ° C: ssa 30 minuuttia, kun sitä käsitellään yleisesti käytetyillä desinfiointiaineilla - 20-30 minuuttia. Virusta sisältävän materiaalin desinfioimiseksi on tarpeen käyttää mykobakteerisidisiä desinfiointiaineita, koska ne ovat tehokkaita mikro-organismeja vastaan, joilla on suurin resistenssi.
HIV-infektion laboratoriodiagnoosi
Tärkein tapa diagnosoida virus ja HIV-infektio on entsyymi-immunoanalyysi. Kuitenkin, johtuen siitä, että gpl20 on rakenteellisia ja antigeeninen samankaltaisuus reseptoreihin ihmisen tiettyjen solujen, mukaan lukien reseptorit, jotka suorittavat liikenteen immunoglobuliinien kautta epiteelisolujen limakalvojen kehossa voi esiintyä liittyvien vasta-aineiden vasta-aineita vastaan gpl20. Tässä tapauksessa IFM: n saattaa olla vääriä positiivisia tuloksia. Näin ollen kaikki positiivisesti reagoivat tutkittujen seerumit altistetaan lisäanalyysille immunoblot-menetelmällä tai western-blottauksella. Tämä menetelmä perustuu elektroforeettisen erotuksen jälkeen tutkittavien vasta-aineiden tunnistamiseen ja sen jälkeiseen testaukseen merkittyjen anti-virus-vasta-aineiden kanssa. Virologisen menetelmän käyttö on vähäistä johtuen viruksen viljelyn monimutkaisuudesta. H9-lymfosyyttien kloonia käytetään viruksen antigeenien saamiseksi - diagnostisten testijärjestelmien tarvittavat komponentit. CDR-menetelmä mahdollistaa viruksen havaitsemisen jo viremian varhaisessa vaiheessa.
HIV-infektion hoito
On tarpeen löytää tai syntetisoida lääkkeitä, jotka estävät tehokkaasti käänteistranskriptaasin (revertase) tai virusproteaasin aktiivisuuden. Ne estäisivät DNA-proviruksen muodostumista ja (tai) estäisivät viruksen solunsisäisen lisääntymisen. Nykyinen hiv-tartunnan saaneiden hoitomuotojen strategia perustuu siihen, että yhdistelmähoitoa estävät virusproteaasit (toinen lääkkeistä) ja reversio (2 erilaista lääkettä), yhdistetty (kolminkertainen) hoito. Venäjällä suositellaan kahden home-lääkkeen käyttöä: fosfatsidia ja crixiaania, joka erityisesti estää HIV-lisääntymisen alkion ja myöhäisten lisääntymisvaiheiden aikana, erityisesti vähentämällä atsidotiimiinin aktiivisuutta, HIV-infektoituneiden ihmisten hoidossa.
Erityisen ennaltaehkäisyn ongelma on tarve luoda rokote, joka varmistaisi tehokkaan soluvälitteisen immuniteetin muodostumisen viruskohtaisten sytotoksisten lymfosyyttien perusteella ilman mitään merkittävää vasta-aineiden tuottamista. Tällainen immuniteetti tarjoaa Thl-apua. On mahdollista, että vasta-aineet, mukaan lukien viruksen neutralointi, ovat paitsi tehottomia HIV-infektion estämisessä, mutta korkealla tasolla ne suppressoivat soluvälitteisen immuniteetin. Siksi anti-HIV-rokotteen tulisi ensisijaisesti täyttää kaksi perusvaatimusta: a) oltava täysin turvallinen ja b) stimuloimaan T-sytotoksisten lymfosyyttien toimintaa. Tutkitaan tapettujen (inaktivoitujen) virusten ja yksittäisten antigeenien, joilla on suuret suojaavat ominaisuudet, tehokkuutta eri rokotteiden muunnoksissa. Tällaisia antigeenejä voidaan joko eristää virioista itse tai syntetisoida kemiallisesti. Geenitekniikan menetelmiin perustuvaa rokotetta ehdotetaan. Se on rekombinantti vaccinia-virus, joka kuljettaa HIV-geenejä, jotka ovat vastuussa sellaisten antigeenien synteesistä, joilla on voimakkaat immunogeeniset ominaisuudet. Päätös näiden rokotteiden tehokkuudesta vie kauan aikaa HIV-infektion inkubaatioajan ja patogeenin suuren vaihtelun vuoksi. Hyvin tehokkaan HIV-rokotteen luominen on kiireellinen perusongelma.