Lääketieteen asiantuntija

Uudet julkaisut
aivolisäke
Viimeksi tarkistettu: 04.07.2025

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
Aivolisäke (hypophysis, s.glandula pituitaria) sijaitsee kiilaluun turkkilaisen aivolisäkkeen kuopassa ja erotetaan kallonontelosta aivojen kovakalvon ulokkeen kautta muodostaen kallon pallean. Pallean aukon kautta aivolisäke on yhteydessä diencephalonin hypotalamuksen infundibulumiin. Aivolisäkkeen poikittaismitta on 10–17 mm, anteroposterior 5–15 mm ja vertikaalinen 5–10 mm. Aivolisäkkeen massa miehillä on noin 0,5 g ja naisilla 0,6 g. Aivolisäkettä peittää ulkopuolelta kapseli.
Aivolisäkkeen kahden eri alkeen kehityksen mukaisesti elimessä erotetaan kaksi lohkoa - etu- ja takalohko. Adenohypofyysi eli etulohko (adenohypophysis, s.lobus anterior) on suurempi ja muodostaa 70-80% aivolisäkkeen kokonaismassasta. Se on tiheämpi kuin takalohko. Etulohkossa erotetaan distaalinen osa (pars distalis), joka sijaitsee aivolisäkkeen etuosassa, väliosa (pars intermedia), joka sijaitsee takalohkon rajalla, ja mukulamainen osa (pars tuberalis), joka ulottuu ylöspäin ja yhdistyy hypotalamuksen infundibulumiin. Verisuonten runsauden vuoksi etulohkolla on vaaleankeltainen väri punertavalla sävyllä. Aivolisäkkeen etulohkon parenkyymiä edustavat useat rauhassolutyypit, joiden säikeiden välissä sijaitsevat sinimuotoiset verisuonet. Puolet (50 %) adenohypofyysisoluista on kromofiilisiä adenosyyttejä, joiden sytoplasmassa on hienorakeisia jyviä, jotka värjäytyvät hyvin kromisuoloilla. Näitä ovat asidofiiliset adenosyytit (40 % kaikista adenohypofyysisoluista) ja basofiiliset adenosyytit (10 %). Basofiilisiin adenosyytteihin kuuluvat gonadotrooppiset, kortikotrooppiset ja tyreotrooppiset endokrinosyytit. Kromofobiset adenosyytit ovat pieniä, niillä on suuri tuma ja pieni määrä sytoplasmaa. Näitä soluja pidetään kromofiilisten adenosyyttien esiasteina. Loput 50 % adenohypofyysisoluista ovat kromofobisia adenosyyttejä.
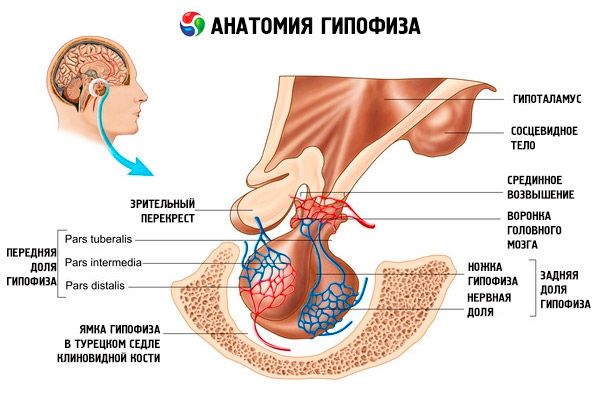
Neurohypofyysi eli takalohko (neurohypofyysi, s.lobus posterior) koostuu hermolohkosta (lobus nervosus), joka sijaitsee aivolisäkkeen takaosassa kuoppaan, ja suppilosta (infundibulum), joka sijaitsee adenohypofyysin tuberkuloosiosan takana. Aivolisäkkeen takalohkon muodostavat neurogliasolut (aivolisäkesolut), hypotalamuksen neurohypofyysiin kulkevat hermokuidut ja neurohypofyysisolut.
Aivolisäke on hermokuitujen (reittien) ja verisuonten kautta toiminnallisesti yhteydessä väliaivoissa sijaitsevaan hypotalamukseen, joka puolestaan säätelee aivolisäkkeen toimintaa. Aivolisäkettä ja hypotalamusta yhdessä neuroendokriinisten, verisuoni- ja hermoyhteyksiensä kanssa pidetään yleensä hypotalamus-aivolisäke-järjestelmänä.
Aivolisäkkeen etu- ja takalohkon hormonit vaikuttavat moniin kehon toimintoihin, pääasiassa muiden umpieritysrauhasten kautta. Aivolisäkkeen etulohkossa asidofiiliset adenosyytit (alfa-solut) tuottavat somatotrooppista hormonia (kasvuhormonia), joka osallistuu nuorten organismien kasvu- ja kehitysprosessien säätelyyn. Kortikotrooppiset endokrinosyytit erittävät adrenokortikotrooppista hormonia (ACTH), joka stimuloi steroidihormonien eritystä lisämunuaisissa. Tyreotrooppiset endokrinosyytit erittävät tyreotrooppista hormonia (TSH), joka vaikuttaa kilpirauhasen kehitykseen ja aktivoi sen hormonien tuotannon. Gonadotrooppiset hormonit: follikkelia stimuloiva (FSH), luteinisoiva (LH) ja prolaktiini - vaikuttavat elimistön sukupuolikypsymiseen, säätelevät ja stimuloivat follikkelien kehitystä munasarjoissa, ovulaatiota, rintarauhasten kasvua ja maidontuotantoa naisilla sekä spermatogeneesiä miehillä. Näitä hormoneja tuottavat basofiiliset adenosyytit (beeta-solut). Myös aivolisäkkeen lipotrooppisia tekijöitä erittyy tänne, jotka vaikuttavat rasvojen mobilisoitumiseen ja hyödyntämiseen kehossa. Etuaivolohkon väliosassa muodostuu melanosyyttejä stimuloivaa hormonia, joka säätelee pigmenttien - melaniinien - muodostumista kehossa.
Hypotalamuksen supraoptisen ja paraventrikulaarisen tumakkeen neurosekretoriset solut tuottavat vasopressiinia ja oksitosiinia. Nämä hormonit kuljetetaan aivolisäkkeen takalohkon soluihin hypotalamus-aivolisäke-järjestelmän muodostavia aksoneita pitkin. Aivolisäkkeen takalohkosta nämä aineet pääsevät verenkiertoon. Vasopressiini-hormonilla on vasokonstriktiivinen ja antidiureettinen vaikutus, minkä vuoksi sitä kutsutaan myös antidiureettiseksi hormoniksi (ADH). Oksitosiinilla on stimuloiva vaikutus kohdun lihasten supistuvuuteen, se lisää maidoneritystä imettävästä rintarauhasesta, estää keltarauhasen kehitystä ja toimintaa sekä vaikuttaa ruoansulatuskanavan sileiden (juovaisten) lihasten sävyn muutoksiin.
Aivolisäkkeen kehitys
Aivolisäkkeen etulohko kehittyy suulahden takaseinämän epiteelistä rengasmaisena kasvuna (Rathken pussi). Tämä ulkonema kasvaa kohti tulevan kolmannen kammion pohjaa. Sitä kohti, toisen aivorakkulan (tulevan kolmannen kammion pohjan) alapinnalta, kasvaa haarake, josta kehittyy infundibulumin harmaa tuberkula ja aivolisäkkeen takalohko.
Aivolisäkkeen verisuonet ja hermot
Aivokalvon valtimoiden sisäisistä kaulavaltimoista ja verisuonista aivolisäkkeen ylempi ja alempi valtimo johtavat aivolisäkkeeseen. Aivolisäkkeen ylemmät valtimot johtavat hypotalamuksen harmaaseen tumakkeeseen ja infundibulumiin, anastomoosittuvat tässä toisiinsa ja muodostavat aivokudokseen tunkeutuvia kapillaareja - primaarista hemokapillaariverkostoa. Tämän verkon pitkistä ja lyhyistä silmukoista muodostuvat porttilaskimot, jotka johtavat aivolisäkkeen etulohkoon. Aivolisäkkeen etulohkon parenkyymissä nämä laskimot hajoavat leveiksi sinusoidaalisiksi kapillaareiksi muodostaen sekundaarisen hemokapillaariverkoston. Aivolisäkkeen takalohko saa verta pääasiassa aivolisäkkeen alemman valtimon kautta. Ylemmän ja alemman aivolisäkkeen valtimoiden välillä on pitkiä valtimoanastomoosiyhteyksiä. Laskimoveren ulosvirtaus sekundaarisesta hemokapillaariverkostosta tapahtuu laskimojärjestelmän kautta, joka virtaa aivojen kovakalvon kavernoottisiin ja kavernoosien välisiin poskionteloihin.
Sympaattiset kuidut, jotka tulevat elimeen valtimoiden mukana, osallistuvat aivolisäkkeen hermotukseen. Postganglioniset sympaattiset hermokuidut lähtevät sisäisen kaulavaltimon hermopunoksesta. Lisäksi aivolisäkkeen takalohkossa on lukuisia hypotalamuksen ytimissä sijaitsevien neurosekretoristen solujen prosessejen päätteitä.
Aivolisäkkeen ikään liittyvät ominaisuudet
Vastasyntyneiden aivolisäkkeen keskimääräinen paino on 0,12 g. Elimen paino kaksinkertaistuu 10 vuoden ikään mennessä ja kolminkertaistuu 15 vuoden ikään mennessä. 20 vuoden ikään mennessä aivolisäkkeen paino saavuttaa maksimiarvonsa (530–560 mg) ja pysyy lähes muuttumattomana seuraavien ikäkausien aikana. 60 vuoden jälkeen tämän umpieritysrauhasen painossa havaitaan lievää laskua.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Aivolisäkkeen hormonit
Hermoston ja hormonaalisen säätelyn yhtenäisyys kehossa varmistetaan aivolisäkkeen ja hypotalamuksen läheisellä anatomisella ja toiminnallisella yhteydellä. Tämä kompleksi määrittää koko hormonaalisen järjestelmän tilan ja toiminnan.
Tärkein umpieritysrauhanen, joka tuottaa useita peptidihormoneja, jotka säätelevät suoraan ääreisrauhasten toimintaa, on aivolisäke. Se on punertavanharmaa pavunmuotoinen muodostuma, jota peittää 0,5–0,6 g painava kuitukapseli. Se vaihtelee hieman henkilön sukupuolen ja iän mukaan. Aivolisäkkeen yleisesti hyväksytty jako kahteen lohkoon, jotka eroavat toisistaan kehityksen, rakenteen ja toiminnan suhteen, on säilynyt: etummainen distaalinen lohko eli adenohypofyysi ja takimmainen neurohypofyysi. Ensimmäinen muodostaa noin 70 % lohkon kokonaismassasta ja se jaetaan tavanomaisesti distaaliseen, infundibulaariseen ja väliosaan, toinen takimmaiseen osaan eli lohkoon ja aivolisäkkeen varteen. Aivolisäke sijaitsee kiilaluun turkkilaisen kuopan aivolisäkkeen kuoppaan ja on yhteydessä aivoihin varren kautta. Etummaisen lohkon yläosaa peittää näköhermon kiasma ja näköradat. Aivolisäkkeen verenkierto on erittäin runsasta ja sitä hoitavat sisäisen kaulavaltimon haarat (aivolisäkkeen ylempi ja alempi valtimo) sekä aivojen valtimokehän haarat. Ylemmät aivolisäkkeen valtimot osallistuvat adenohypofyysin verenkiertoon ja alemmat neurohypofyysin, koskettaen hypotalamuksen suurisoluisten tumakkeiden aksonien neurosekretorisia päitä. Ensimmäiset valtimot kulkevat hypotalamuksen keskieminenssiin, jossa ne jakautuvat kapillaariverkostoksi (primaarinen kapillaaripunos). Nämä kapillaarit (joihin hypotalamuksen keskieminenssin pienten neurosekretoristen solujen aksonien päät ovat kosketuksissa) kerääntyvät porttilaskimoihin, jotka laskeutuvat aivolisäkkeen vartta pitkin adenohypofyysin parenkyymiin, jossa ne jälleen jakautuvat sinusomaalisten kapillaarien verkostoksi (sekundaarinen kapillaaripunos). Näin veri, joka on aiemmin kulkenut hypotalamuksen keskieminenssin läpi, jossa se rikastuu hypotalamuksen adenohypofysotrooppisilla hormoneilla (vapauttavilla hormoneilla), saapuu adenohypofyysiin.
Veren ulosvirtaus, joka on kyllästetty adenohypofyysihormoneilla lukuisista sekundaarisen plexuksen kapillaareista, tapahtuu laskimojärjestelmän kautta, joka puolestaan virtaa kovakalvon laskimoiden poskionteloihin ja sitten yleiseen verenkiertoon. Näin ollen aivolisäkkeen portaalijärjestelmä, jossa veren virtaussuunta on laskeva hypotalamuksesta, on morfofunktionaalinen osa adenohypofyysin trooppisten toimintojen neurohumoraalisen säätelyn monimutkaista mekanismia.
Aivolisäkettä hermottavat sympaattiset kuidut, jotka seuraavat aivolisäkkeen valtimoita. Ne saavat alkunsa postganglionaalisista kuiduista, jotka kulkevat sisäisen kaulavaltimonpunoksen läpi ja ovat yhteydessä ylempään kaulaganglioon. Hypotalamuksella ei ole suoraa hermotusta adenohypofyysiin. Takalohko saa hermokuituja hypotalamuksen neurosekretorisista ytimistä.
Adenohypofyysi on histologiselta rakenteeltaan hyvin monimutkainen muodostuma. Siinä on kahdenlaisia rauhassoluja - kromofobisia ja kromofiilisiä. Jälkimmäiset puolestaan jaetaan asidofiilisiin ja basofiilisiin (aivolisäkkeen yksityiskohtainen histologinen kuvaus on annettu käyttöohjeen vastaavassa osiossa). On kuitenkin huomattava, että adenohypofyysin parenkyymin tuottamat hormonit ovat kemialliselta luonteeltaan jossain määrin erilaisia jälkimmäisten monimuotoisuuden vuoksi, ja erittävien solujen hienorakenteen on vastattava kunkin solujen biosynteesin ominaisuuksia. Mutta joskus adenohypofyysissä voidaan havaita rauhassolujen siirtymämuotoja, jotka kykenevät tuottamaan useita hormoneja. On näyttöä siitä, että adenohypofyysin rauhassolujen tyyppi ei aina ole geneettisesti määrätty.
Turkkilaisen kalvon pallean alla on aivolisäkkeen etulohkon suppilomainen osa. Se ympäröi aivolisäkkeen vartta ja on kosketuksissa harmaaseen kyhmyyn. Tälle adenohypofyysin osalle on ominaista epiteelisolujen läsnäolo ja runsas verenkierto. Se on myös hormonaalisesti aktiivinen.
Aivolisäkkeen väliosa (keskimmäinen) koostuu useista kerroksista suuria eritysaktiivisia basofiilisiä soluja.
Aivolisäke suorittaa useita toimintoja hormoniensa kautta. Sen etulohko tuottaa adrenokortikotrooppista (ACTH), kilpirauhasta stimuloivaa (TSH), follikkelia stimuloivaa (FSH), luteinisoivaa (LH), lipotrooppisia hormoneja sekä kasvuhormonia - somatotrooppista (STO) ja prolaktiinia. Välilohkossa syntetisoidaan melanosyyttejä stimuloivaa hormonia (MSH), ja takalohkossa kertyvät vasopressiini ja oksitosiini.
ACTH
Aivolisäkkeen hormonit ovat ryhmä proteiini- ja peptidihormoneja sekä glykoproteiineja. Aivolisäkkeen etulohkon hormoneista ACTH:ta on tutkittu parhaiten. Sitä tuottavat basofiiliset solut. Sen tärkein fysiologinen tehtävä on stimuloida steroidihormonien biosynteesiä ja eritystä lisämunuaisen kuoressa. ACTH:lla on myös melanosyyttejä stimuloivaa ja lipotrooppista aktiivisuutta. Vuonna 1953 se eristettiin puhtaassa muodossa. Myöhemmin sen kemiallinen rakenne, joka koostuu 39 aminohappotähteestä ihmisillä ja useilla nisäkkäillä, määritettiin. ACTH:lla ei ole lajispesifisyyttä. Tällä hetkellä on suoritettu sekä itse hormonin että sen molekyylin eri fragmenttien kemiallinen synteesi, jotka ovat aktiivisempia kuin luonnolliset hormonit. Hormonin rakenteessa on kaksi peptidiketjun osaa, joista toinen varmistaa ACTH:n havaitsemisen ja sitoutumisen reseptoriin, ja toinen tarjoaa biologisen vaikutuksen. Se sitoutuu ilmeisesti ACTH-reseptoriin hormonin ja reseptorin sähkövarausten vuorovaikutuksen kautta. ACTH:n biologisen efektorin roolin suorittaa 4-10-molekyylin fragmentti (Met-Glu-His-Phen-Arg-Tri-Tri).
ACTH:n melanosyyttejä stimuloiva vaikutus johtuu molekyylin N-terminaalisesta alueesta, joka koostuu 13 aminohappotähteestä ja toistaa alfa-melanosyyttejä stimuloivan hormonin rakennetta. Tämä alue sisältää myös heptapeptidin, jota esiintyy muissa aivolisäkkeen hormoneissa ja jolla on joitakin adrenokortikotrooppisia, melanosyyttejä stimuloivia ja lipotrooppisia vaikutuksia.
ACTH:n vaikutuksen avainhetkenä pidetään proteiinikinaasientsyymin aktivaatiota sytoplasmassa cAMP:n osallistuessa. Fosforyloitu proteiinikinaasi aktivoi esteraasientsyymin, joka muuntaa kolesteroliesterit vapaaksi aineeksi rasvapisaroissa. Ribosomien fosforylaation seurauksena sytoplasmassa syntetisoitu proteiini stimuloi vapaan kolesterolin sitoutumista sytokromi P-450:een ja sen siirtymistä lipidipisaroista mitokondrioihin, joissa ovat läsnä kaikki entsyymit, jotka varmistavat kolesterolin muuntumisen kortikosteroideiksi.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Kilpirauhasta stimuloiva hormoni
TSH - tyreotropiini - kilpirauhasen kehityksen ja toiminnan, kilpirauhashormonien synteesin ja erityksen prosessien pääasiallinen säätelijä. Tämä monimutkainen proteiini - glykoproteiini - koostuu alfa- ja beeta-alayksiköistä. Ensimmäisen alayksikön rakenne vastaa luteinisoivaa hormonia (TSH) ja sen alfa-alayksikköä. Lisäksi se on suurelta osin sama eri eläinlajeilla. Ihmisen TSH:n beeta-alayksikön aminohappotähteiden järjestys on selvitetty ja se koostuu 119 aminohappotähteestä. Voidaan havaita, että ihmisen ja naudan TSH:n beeta-alayksiköt ovat monella tapaa samankaltaisia. Glykoproteiinihormonien biologiset ominaisuudet ja biologisen aktiivisuuden luonne määräytyvät beeta-alayksikön perusteella. Se varmistaa myös hormonin vuorovaikutuksen reseptorien kanssa eri kohde-elimissä. Useimmilla eläimillä beeta-alayksikkö osoittaa kuitenkin spesifistä aktiivisuutta vasta yhdistyttyään alfa-alayksikköön, joka toimii eräänlaisena hormonin aktivaattorina. Jälkimmäinen indusoi yhtä todennäköisesti luteinisoivaa, follikkelia stimuloivaa ja tyreotrooppista aktiivisuutta, jotka määräytyvät beeta-alayksikön ominaisuuksien perusteella. Löydetty samankaltaisuus antaa meille mahdollisuuden päätellä, että nämä hormonit syntyivät evoluutioprosessissa yhdestä yhteisestä edeltäjästä, beeta-alayksikkö määrittää myös hormonien immunologiset ominaisuudet. Oletetaan, että alfa-alayksikkö suojaa beeta-alayksikköä proteolyyttisten entsyymien vaikutukselta ja helpottaa myös sen kulkeutumista aivolisäkkeestä ääreishermoston kohdeelimiin.
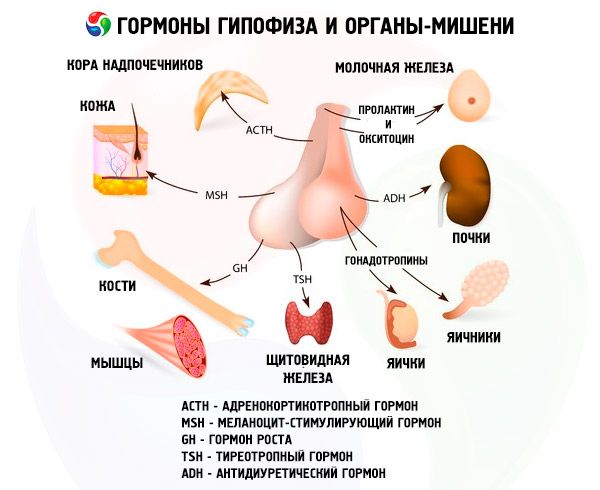
Gonadotrooppiset hormonit
Gonadotropiineja esiintyy elimistössä LH:n ja FSH:n muodossa. Näiden hormonien toiminnallinen tarkoitus rajoittuu yleensä lisääntymisprosessien varmistamiseen molemmilla sukupuolilla. Ne, kuten TSH, ovat monimutkaisia proteiineja - glykoproteiineja. FSH indusoi follikkelien kypsymistä naisten munasarjoissa ja stimuloi spermatogeneesiä miehillä. LH aiheuttaa naisilla follikkelien repeämisen ja keltarauhasen muodostumisen sekä stimuloi estrogeenien ja progesteronin eritystä. Miehillä sama hormoni kiihdyttää kudoskudoksen kehitystä ja androgeenien eritystä. Gonadotropiinien vaikutukset ovat toisistaan riippuvaisia ja tapahtuvat synkronisesti.
Gonadotropiinien erityksen dynamiikka naisilla muuttuu kuukautiskierron aikana, ja sitä on tutkittu riittävän yksityiskohtaisesti. Kierron preovulaatiovaiheessa (follikulaarisessa) LH-pitoisuus on melko alhainen, kun taas FSH-pitoisuus on koholla. Follikkelin kypsyessä estradiolin eritys lisääntyy, mikä edistää aivolisäkkeen gonadotropiinien tuotannon lisääntymistä ja sekä LH- että FSH-syklien käynnistymistä eli sukupuolisteroidit stimuloivat gonadotropiinien eritystä.
LH:n rakenne on tällä hetkellä määritetty. Kuten TSH, se koostuu kahdesta alayksiköstä: a:sta ja b:stä. LH:n alfa-alayksikön rakenne eri eläinlajeilla on suurelta osin samanlainen, se vastaa TSH:n alfa-alayksikön rakennetta.
LH:n beeta-alayksikön rakenne eroaa merkittävästi TSH:n beeta-alayksikön rakenteesta, vaikka sillä on neljä identtistä peptidiketjun osaa, jotka koostuvat 4-5 aminohappotähteestä. TSH:ssa ne sijaitsevat asemissa 27-31, 51-54, 65-68 ja 78-83. Koska LH:n ja TSH:n beeta-alayksikkö määrää hormonien spesifisen biologisen aktiivisuuden, voidaan olettaa, että LH:n ja TSH:n rakenteessa olevat homologiset osat varmistavat beeta-alayksiköiden yhteyden alfa-alayksikköön, ja rakenteeltaan erilaiset osat vastaavat hormonien biologisen aktiivisuuden spesifisyydestä.
Natiivi LH on erittäin stabiili proteolyyttisten entsyymien vaikutukselle, mutta kymotrypsiini pilkkoo beeta-alayksikön nopeasti ja entsyymi hydrolysoi vaikeasti α-alayksikköä, eli sillä on suojaava rooli estäen kymotrypsiiniä pääsemästä peptidisidoksiin.
FSH:n kemiallisen rakenteen osalta tutkijat eivät ole vielä saaneet lopullisia tuloksia. Kuten LH, FSH koostuu kahdesta alayksiköstä, mutta FSH:n beeta-alayksikkö eroaa LH:n beeta-alayksiköstä.
Prolaktiini
Toinen hormoni, prolaktiini (laktogeeninen hormoni), on aktiivinen lisääntymisprosesseissa. Prolaktiinin tärkeimmät fysiologiset ominaisuudet nisäkkäillä ilmenevät rintarauhasten ja imetyksen kehityksen stimuloinnissa, talirauhasten ja sisäelinten kasvussa. Se edistää steroidien vaikutusta toissijaisiin sukupuoliominaisuuksiin uroksilla, stimuloi keltarauhasen eritystoimintaa hiirillä ja rotilla sekä osallistuu rasva-aineenvaihdunnan säätelyyn. Prolaktiiniin on viime vuosina kiinnitetty paljon huomiota äidin käyttäytymisen säätelijänä; tällainen polyfunktionaalisuus selittyy sen evolutiivisella kehityksellä. Se on yksi muinaisista aivolisäkkeen hormoneista ja sitä esiintyy jopa sammakkoeläimillä. Tällä hetkellä prolaktiinin rakenne joillakin nisäkäslajeilla on täysin selvitetty. Tutkijat ovat kuitenkin äskettäin ilmaisseet epäilyksensä tällaisen hormonin olemassaolosta ihmisillä. Monet uskoivat, että sen tehtävän suorittaa kasvuhormoni. Nyt on saatu vakuuttavia todisteita prolaktiinin esiintymisestä ihmisillä ja sen rakenne on osittain selvitetty. Prolaktiinireseptorit sitoutuvat aktiivisesti kasvuhormoniin ja istukan laktogeeniin, mikä viittaa näiden kolmen hormonin yhteen vaikutusmekanismiin.
Somatotropiini
Kasvuhormonilla, somatotropiinilla, on vieläkin laajempi vaikutuskirjo kuin prolaktiinilla. Kuten prolaktiini, sitä tuottavat adenohypofyysin asidofiiliset solut. STH stimuloi luuston kasvua, aktivoi proteiinisynteesiä, sillä on rasvaa mobilisoiva vaikutus ja se edistää kehon koon kasvua. Lisäksi se koordinoi aineenvaihduntaprosesseja.
Hormonin osallistuminen jälkimmäiseen vahvistetaan sillä, että aivolisäkkeen eritys lisääntyy jyrkästi esimerkiksi verensokeritason laskiessa.
Tämän ihmishormonin kemiallinen rakenne on nyt täysin selvillä – 191 aminohappotähtettä. Sen ensisijainen rakenne on samanlainen kuin istukkahormonin somatomammotropiinin tai istukan laktogeenin. Nämä tiedot osoittavat näiden kahden hormonin merkittävää evolutiivista läheisyyttä, vaikka niiden biologinen aktiivisuus eroaakin.
On tarpeen korostaa kyseisen hormonin korkeaa lajispesifisyyttä - esimerkiksi eläinperäinen kasvuhormoni on inaktiivinen ihmisillä. Tämä selittyy sekä ihmisen ja eläimen kasvuhormonin reseptorien välisellä reaktiolla että itse hormonin rakenteella. Tällä hetkellä on käynnissä tutkimuksia, joilla tunnistetaan aktiivisia keskuksia kasvuhormonin monimutkaisessa rakenteessa, joilla on biologista aktiivisuutta. Tutkitaan myös molekyylin yksittäisiä fragmentteja, joilla on muita ominaisuuksia. Esimerkiksi ihmisen kasvuhormonin pepsiinillä hydrolyysin jälkeen eristettiin peptidi, joka koostui 14 aminohappotähteestä ja vastasi molekyylin osaa 31-44. Sillä ei ollut kasvuvaikutusta, mutta se ylitti merkittävästi natiivin hormonin lipotrooppisessa aktiivisuudessa. Ihmisen kasvuhormonilla, toisin kuin vastaavalla eläinten hormonilla, on merkittävä laktogeeninen aktiivisuus.
Adenohypofyysi syntetisoi monia peptidi- ja proteiiniaineita, joilla on rasvaa mobilisoiva vaikutus, ja aivolisäkkeen trooppisilla hormoneilla - ACTH, STH, TSH ja muilla - on lipotrooppinen vaikutus. Viime vuosina on korostettu erityisesti beeta- ja γ-lipotrooppisia hormoneja (LPG). Beeta-LPG:n biologisia ominaisuuksia on tutkittu yksityiskohtaisemmin; lipotrooppisen aktiivisuuden lisäksi sillä on myös melanosyyttejä stimuloiva, kortikotropiinia stimuloiva ja hypokalseminen vaikutus, ja se tuottaa myös insuliinin kaltaisen vaikutuksen.
Tällä hetkellä lampaiden LPG:n (90 aminohappotähtettä), sikojen ja nautojen lipotrooppisten hormonien, primaarirakenne on selvitetty. Tällä hormonilla on lajispesifisyyttä, vaikka beeta-LPG:n keskusalueen rakenne on sama eri lajeilla. Se määrittää hormonin biologiset ominaisuudet. Yksi tämän alueen fragmenteista löytyy alfa-MSH:n, beeta-MSH:n, ACTH:n ja beeta-LPG:n rakenteesta. On ehdotettu, että nämä hormonit ovat syntyneet samasta esiasteesta evoluutioprosessissa. γ-LPG:llä on heikompi lipotrooppinen aktiivisuus kuin beeta-LPG:llä.
Melanosyyttejä stimuloiva hormoni
Tämä aivolisäkkeen välilohkossa syntetisoitu hormoni stimuloi ihon pigmentti melaniinin biosynteesiä biologisessa tehtävässään ja edistää pigmenttisolujen, melanosyyttien, koon ja lukumäärän kasvua sammakkoeläinten ihossa. Näitä MSH:n ominaisuuksia käytetään hormonin biologisissa testeissä. Hormonia on kahdenlaisia: alfa- ja beeta-MSH. On osoitettu, että alfa-MSH:lla ei ole lajispesifisyyttä ja sillä on sama kemiallinen rakenne kaikilla nisäkkäillä. Sen molekyyli on peptidiketju, joka koostuu 13 aminohappotähteestä. Beeta-MSH:lla sitä vastoin on lajispesifisyys, ja sen rakenne vaihtelee eri eläimillä. Useimmilla nisäkkäillä beeta-MSH-molekyyli koostuu 18 aminohappotähteestä, ja vain ihmisillä se on pidennetty aminohappopäästä neljällä aminohappotähteellä. On huomattava, että alfa-MSH:lla on jonkin verran adrenokortikotrooppista aktiivisuutta, ja sen vaikutus eläinten ja ihmisten käyttäytymiseen on nyt todistettu.
Oksitosiini ja vasopressiini
Hypotalamuksessa syntetisoituvat vasopressiini ja oksitosiini kerääntyvät aivolisäkkeen takalohkoon: vasopressiini supraoptisen tumakkeen hermosoluihin ja oksitosiini paraventrikulaariseen tumakkeeseen. Sitten ne siirtyvät aivolisäkkeeseen. On korostettava, että vasopressiinihormonin esiaste syntetisoidaan ensin hypotalamuksessa. Samalla siellä tuotetaan neurofysiini tyypit 1 ja 2. Ensimmäinen sitoutuu oksitosiiniin ja toinen vasopressiiniin. Nämä kompleksit kulkeutuvat neurosekretoristen rakeiden muodossa sytoplasmassa aksonia pitkin ja saavuttavat aivolisäkkeen takalohkon, jossa hermokuidut päättyvät verisuonen seinämään ja rakeiden sisältö pääsee vereen. Vasopressiini ja oksitosiini ovat ensimmäiset aivolisäkehormonit, joilla on täysin vakiintunut aminohapposekvenssi. Kemialliselta rakenteeltaan ne ovat nonapeptidejä, joissa on yksi disulfidisilta.
Tarkasteltavilla hormoneilla on erilaisia biologisia vaikutuksia: ne stimuloivat veden ja suolojen kuljetusta kalvojen läpi, niillä on vasopressorinen vaikutus, ne tehostavat kohdun sileiden lihasten supistumista synnytyksen aikana ja lisäävät rintarauhasten eritystä. On huomattava, että vasopressiinilla on suurempi antidiureettinen aktiivisuus kuin oksitosiinilla, kun taas jälkimmäisellä on voimakkaampi vaikutus kohtuun ja rintarauhasiin. Vasopressiinin erityksen pääasiallinen säätelijä on veden kulutus; munuaistiehyissä se sitoutuu sytoplasmakalvojen reseptoreihin, jolloin niissä aktivoituu adenylaattisyklaasientsyymi. Molekyylin eri osat vastaavat hormonin sitoutumisesta reseptoriin ja biologisesta vaikutuksesta.
Aivolisäke, joka on hypotalamuksen kautta yhteydessä koko hermostoon, yhdistää umpieritysjärjestelmän toiminnalliseksi kokonaisuudeksi ja osallistuu elimistön sisäisen ympäristön pysyvyyden (homeostaasin) varmistamiseen. Umpieritysjärjestelmän sisällä homeostaattinen säätely tapahtuu aivolisäkkeen etulohkon ja "kohde"rauhasten (kilpirauhanen, lisämunuaisen kuori, sukurauhaset) välisen palautteen periaatteella. "Kohde"rauhasen tuottaman hormonin liikatuotanto estää ja sen puutos stimuloi vastaavan trooppisen hormonin eritystä ja vapautumista. Hypotalamus kuuluu takaisinkytkentäjärjestelmään. Siinä sijaitsevat "kohde"rauhasten hormoneille herkät reseptorivyöhykkeet. Sitoutumalla spesifisesti veressä kiertäviin hormoneihin ja muuttamalla vastetta hormonipitoisuudesta riippuen hypotalamuksen reseptorit välittävät vaikutuksensa vastaaviin hypotalamuksen keskuksiin, jotka koordinoivat adenohypofyysin työtä vapauttaen hypotalamuksen adenohypofysiotrooppisia hormoneja. Siksi hypotalamusta tulisi pitää neuroendokriinisenä aivona.