Lääketieteen asiantuntija

Uudet julkaisut
Munasarja
Viimeksi tarkistettu: 04.07.2025

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
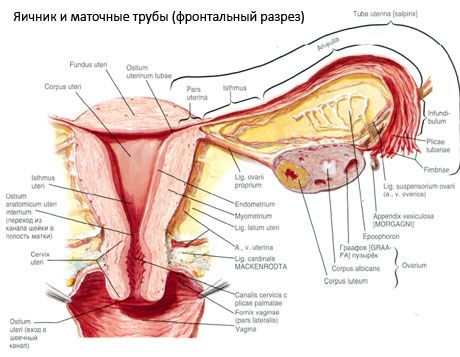
Munasarja (ovarium; kreikaksi oophoron) on parillinen elin, naisen lisääntymisrauhanen, joka sijaitsee lantionontelossa kohdun leveän nivelsiteen takana. Munasarjoissa kehittyvät ja kypsyvät naisen lisääntymissolut (munasolut), ja muodostuu naissukuhormoneja, jotka kulkeutuvat vereen ja imusolmukkeisiin. Munasarja on muodoltaan soikea, hieman litistynyt anteroposteriorisesta suunnasta. Munasarjan väri on vaaleanpunainen. Synnyttäneen naisen munasarjan pinnalla näkyy painaumia ja arpia - jälkiä ovulaatiosta ja keltarauhasen muutoksesta. Munasarja painaa 5-8 g. Munasarjan mitat ovat: pituus 2,5-5,5 cm, leveys 1,5-3,0 cm, paksuus - jopa 2 cm. Munasarjassa on kaksi vapaata pintaa: lantiononteloon päin oleva mediaalinen pinta (facies medialis), osittain munanjohtimen peittämä, ja lateraalinen pinta (facies lateralis), joka on lantion sivuseinän vieressä hieman voimakkaan painauman - munasarjan kuopan - vieressä. Tämä kuoppa sijaitsee ulompien, vatsakalvon peittämien suoliluun verisuonten ja alhaalla olevien kohdun ja sulkuvaltimoiden välisessä kulmassa. Munasarjan takana vastaavan puolen virtsanjohdin kulkee retroperitoneaalisesti ylhäältä alas.
Munasarjan pinnat kulkevat kuperaan vapaaseen (taka) reunaan (margo liber), edessä suoliliepeen reunaan (margo mesovaricus), joka on kiinnitetty vatsakalvon (munasarjan suoliliepeen) lyhyen taitoksen avulla kohdun leveän nivelsiteen takaosaan. Tällä elimen etureunalla on uurteinen painauma - munasarjan hilum (hilum ovarii), jonka kautta valtimo ja hermot tulevat munasarjaan ja laskimot ja imusuonet poistuvat. Munasarjalla on myös kaksi päätä: pyöreä ylempi putkimainen pää (extremitas tubaria), joka on munanjohtimeen päin, ja alempi kohdun pää (extremitas utenna), joka on yhdistetty kohtuun munasarjan nivelsiteen (lig. ovarii proprium) avulla. Tämä noin 6 mm paksuinen pyöreä naru muodostaa nivelsiteen, joka kulkee munasarjan kohdunpuoleisesta päästä kohdun sivukulmaan, joka sijaitsee kohdun leveän nivelsiteen kahden lehden välissä. Munasarjan nivelsidejärjestelmään kuuluu myös munasarjan nivelside suspensorium (lig.suspensorium ovarii), joka on vatsakalvon poimu, joka kulkee pienen lantion seinämästä munasarjaan ja sisältää munasarjan verisuonet ja kuituisten kuitujen kimppuja. Munasarjaa kiinnittää lyhyt suolilieve (mesovarium), joka on vatsakalvon kaksoiskappale, joka kulkee kohdun leveän nivelsiteen takaläppästä munasarjan suolilieveen. Itse munasarjat eivät ole vatsakalvon peitossa. Munanjohtimen suurin munasarjareunus on kiinnittynyt munasarjan putkimaiseen päähän. Munasarjan topografia riippuu kohdun sijainnista ja koosta (raskauden aikana). Munasarjat ovat pienen lantion ontelon erittäin liikkuvia elimiä.
 [
[ Munasarjojen verisuonet ja hermot
Munasarjojen verenkiertoa hoitavat munasarjat aa. et vv. ovaricae et uterinae. Molemmat munasarjavaltimot (aa. ovaricae dextra et sinistra) saavat alkunsa aortan etupinnalta juuri munuaisvaltimoiden alapuolelta; oikea valtimo yleensä lähtee aortasta ja vasen munuaisvaltimosta. Munasarjojen valtimot kulkevat alaspäin ja sivusuunnassa pitkin psoas major -lihaksen etupintaa ja ylittävät edessä olevan virtsanjohtimen (haaroittuen siihen), ulommat suoliluun verisuonet, niiden reunaviivan ja johtavat lantioonteloon, joka sijaitsee tässä munasarjan ripustusligamentissa. Mediaalisessa suunnassa munasarjavaltimo kulkee kohdun leveän nivelsiteen lehtien välistä munanjohtimen alla, haarautuen siihen, ja sitten munasarjan suolilieveeseen; se kulkee munasarjan hilumiin.
Munasarjavaltimon haarat yhtyvät laajasti kohdun valtimon munasarjahaaroihin. Laskimovirtaus munasarjoista tapahtuu pääasiassa munasarjojen laskimopunokseen, joka sijaitsee munasarjojen kukkulan alueella. Täältä veri virtaa kahteen suuntaan: kohdun ja munasarjojen laskimoiden kautta. Oikeassa munasarjalaskimossa on läppiä, ja se virtaa alaonttolaskimoon. Vasen munasarjalaskimo virtaa vasempaan munuaislaskimoon, vaikka siinä ei ole läppiä.
Munasarjoista imuneste poistuu imusuonten kautta, erityisesti runsaasti elinportin alueella, jossa erottuu subovarianinen imusuonten punos. Sitten imuneste poistuu munasarjojen imusuonia pitkin paraaorttisiin imusolmukkeisiin.
Munasarjojen hermotus
Sympaattinen - keliakian (aurinkohermon), ylemmän mesenteerisen ja hypogastrisen hermopunoksen postganglionaalisten kuidujen tarjoama; parasympaattinen - viskeraalisten sakraalihermojen kautta.
Munasarjan rakenne
Munasarjan pintaa peittää yksikerroksinen alkioepiteeli. Sen alla on tiheä sidekudosproteiinikuori (tunica albuginea). Munasarjan sidekudos muodostaa sen strooman (stroma ovarii), joka on runsas elastisista kuiduista. Munasarjan aines, sen parenkyymi, jaetaan ulko- ja sisäkerrokseen. Sisäkerros, joka sijaitsee munasarjan keskellä, lähempänä sen porttia, on nimeltään ydin (medulla ovarii). Tässä kerroksessa, löyhässä sidekudoksessa, on lukuisia veri- ja imusuonia sekä hermoja. Munasarjan ulkokerros - kuorikerros (cortex ovarii) - on tiheämpi. Se sisältää paljon sidekudosta, jossa sijaitsevat kypsyvät primaariset munarakkulat (folliculi ovarici primarii), sekundaariset (vesikulaariset) follikkelit (folliculi ovarici secundarii, s.vesiculosi) sekä kypsät follikkelit, Graafin follikkelit (folliculi ovarici maturis) sekä keltaiset ja atreettiset kappaleet.
Jokainen follikkeli sisältää naaraspuolisen lisääntymismunasolun eli oosyytin (ococytus). Munasolu on halkaisijaltaan jopa 150 µm, pyöreä, sisältää tuman ja suuren määrän sytoplasmaa, joka soluorganellien lisäksi sisältää proteiini-lipidi-inkluusiota (keltuainen) ja glykogeenia, jotka ovat välttämättömiä munasolun ravinnolle. Munasolu yleensä käyttää ravintoaineensa loppuun 12–24 tunnin kuluessa ovulaatiosta. Jos hedelmöitystä ei tapahdu, munasolu kuolee.
Munasolua peittää kaksi kalvoa. Sisällä on sytolemma, joka on munasolun sytoplasmakalvo. Sytolemman ulkopuolella on kerros niin kutsuttuja follikkelisoluja, jotka suojaavat munasolua ja suorittavat hormonien muodostavan tehtävän – ne erittävät estrogeeneja.
Kohdun, munanjohtimien ja munasarjojen fysiologisen asennon tarjoavat vatsakalvon, nivelsiteet ja lantion kudoksen yhdistävät ripustus-, kiinnitys- ja tukilaitteet. Ripustuslaite muodostuu parillisista muodostelmista, ja siihen kuuluvat kohdun pyöreät ja leveät nivelsiteet, munasarjojen oikeat nivelsiteet ja ripustusnivelsiteet. Kohdun leveät nivelsiteet, munasarjojen oikeat ja ripustusnivelsiteet pitävät kohdun keskiasennossa. Pyöreät nivelsiteet vetävät kohdun pohjaa eteenpäin ja tarjoavat sen fysiologisen kallistuksen.
Kiinnityslaite varmistaa kohdun sijainnin pienen lantion keskellä ja tekee käytännössä mahdottomaksi sen siirtymisen sivuille, edestakaisin. Mutta koska nivelsidelaite irtoaa kohdusta alaosastaan, kohdun kallistuminen eri suuntiin on mahdollista. Kiinnityslaite sisältää lantion löysässä kudoksessa sijaitsevat nivelsiteet, jotka ulottuvat kohdun alaosasta lantion sivu-, etu- ja takaseiniin: sakromagiset, kardinaali-, uterovesikaaliset ja vesikopubiset nivelsiteet.
Mesovariumin lisäksi erotetaan seuraavat munasarjojen nivelsiteet:
- munasarjan ripustava nivelside, aiemmin infundibulopelvic-nivelside. Se on vatsakalvon poimu, jonka läpi kulkevat munasarjan verisuonet (a. et v. ovarica), imusuonet ja hermot. Se on venytetty lantion sivuseinän, lannekalvon faskian (yhteisen suoliluunvaltimon jakauman alueella ulkoiseen ja sisäiseen) ja munasarjan yläpään (munajohtimien) väliin.
- Munasarjan oikea nivelside kulkee leveän kohdun nivelsiteen lehtien välistä, lähempänä takalehteä, ja yhdistää munasarjan alapään kohdun sivureunaan. Munasarjan oikea nivelside on kiinnittynyt kohtuun munanjohtimen alun ja pyöreän nivelsiteen väliin, jälkimmäisen taakse ja yläpuolelle. Ligamentin paksuudessa kulkee oikeanpuoleiset munasarjat (vr. ovarii), jotka ovat kohdun valtimon päähaarat;
- Apukalvon ja munasarjan välinen nivelside. Ligamentti ulottuu umpilisäkkeen suolesta kohdun oikeaan munasarjaan tai leveään nivelsideeseen vatsakalvon poimuna. Ligamentti on epävakaa ja sitä esiintyy 1/2–1/3 naisista.
Tukilaitetta edustavat lantionpohjan lihakset ja faskiat, jotka on jaettu alempaan, keskimmäiseen ja ylempään (sisäiseen) kerrokseen.
Voimakkain on ylempi (sisempi) lihaskerros, jota edustaa peräaukkoa nostava parillinen lihas. Se koostuu lihaskimpuista, jotka levittäytyvät häntäluusta lantion luihin kolmeen suuntaan (pubococcygeus, iliococcygeus ja ischiococcygeus). Tätä lihaskerrosta kutsutaan myös lantion palleaksi.
Keskimmäinen lihaskerros sijaitsee symphysis-, häpy- ja iskialuiden välissä. Keskimmäinen lihaskerros - urogenitaalinen pallea - sijaitsee lantion ulostuloaukon etuosassa, jonka läpi virtsaputki ja emätin kulkevat. Sen arkkien välissä etuosassa ovat lihaskimput, jotka muodostavat virtsaputken ulomman sulkijalihaksen, ja takaosassa ovat lihaskimput, jotka kulkevat poikittaissuunnassa - välilihan syvä poikittainen lihas.
Lantionpohjan lihasten alempi (ulompi) kerros koostuu pinnallisista lihaksista, joiden muoto muistuttaa numeroa 8. Näitä ovat bulbokavernoottinen lihas, ischiokavernoottinen lihas, ulompi peräaukon sulkijalihas ja pinnallinen poikittainen välilihan lihas.
Munasarjojen ontogeneesi
Follikkelien kasvu ja atresia alkavat 20. raskausviikolla, ja synnytykseen mennessä tytön munasarjoissa on jäljellä jopa 2 miljoonaa munasolua. Kuukautisten alkamiseen mennessä niiden määrä laskee 300 tuhanteen. Koko lisääntymiselämän aikana enintään 500 follikkelia saavuttaa kypsyyden ja ovuloi. Follikkelien alkukasvu ei ole riippuvainen FSH-stimulaatiosta, on rajallista ja atresia tapahtuu nopeasti. Uskotaan, että steroidihormonien sijaan paikalliset autokriiniset/parakriiniset peptidit ovat ensisijaisten follikkelien kasvun ja atresian pääasiallinen säätelijä. Uskotaan, että follikkelien kasvua ja atresiaa ei keskeytä mikään fysiologinen prosessi. Tämä prosessi jatkuu kaikissa ikäryhmissä, mukaan lukien kohdunsisäinen kausi ja vaihdevuodet, ja sen keskeyttävät raskaus, ovulaatio ja anovulaatio. Mekanismi, joka laukaisee follikkelien kasvun ja niiden määrän kussakin tietyssä syklissä, on edelleen epäselvä.
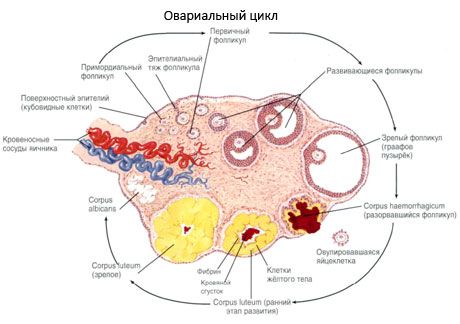
Kehityksensä aikana follikkeli käy läpi useita kehitysvaiheita. Alkusolut lähtevät ruskuaispussin endodermista, allantoisista, ja siirtyvät alkion sukupuolielimille 5–6 raskausviikolla. Nopean mitoottisen jakautumisen seurauksena, joka jatkuu 6–8 raskausviikosta 16–20 raskausviikkoon, alkion munasarjoihin muodostuu jopa 6–7 miljoonaa munasolua, joita ympäröi ohut granulosa-solukerros.
Preantraalfollikkeli - munasolua ympäröi kalvo (Zona pellucida). Munasolua ympäröivät granulosasolut alkavat lisääntyä, niiden kasvu riippuu gonadotropiineista ja korreloi estrogeenitason kanssa. Granulosasolut ovat FSH:n kohde. Preantraalfollikkelivaiheessa granulosasolut kykenevät syntetisoimaan kolmenlaisia steroideja: indusoivat pääasiassa aromataasin, androgeenit estradioliksi muuntavan pääentsyymin, aktiivisuutta. Estradiolin uskotaan kykenevän lisäämään omien reseptoriensa määrää ja siten aiheuttamaan suoran mitogeenisen vaikutuksen granulosasoluihin FSH:sta riippumatta. Sitä pidetään parakriinisena tekijänä, joka tehostaa FSH:n vaikutuksia, mukaan lukien aromatisaatioprosessien aktivointi.
FSH-reseptoreita ilmestyy granulosasolujen kalvoille heti follikkelien kasvun alkaessa. FSH:n väheneminen tai lisääntyminen johtaa sen reseptorien määrän muutokseen. Kasvutekijät säätelevät tätä FSH:n vaikutusta. FSH vaikuttaa G-proteiinin, adenylaattisyklaasijärjestelmän, kautta, ja vaikka steroidogeneesiä follikkeleissa säätelee pääasiassa FSH, tähän prosessiin osallistuu monia tekijöitä: ionikanavia, tyrosiinikinaasireseptoreita ja toisiolähettien fosfolipaasijärjestelmää.
Androgeenien rooli follikkelien varhaisessa kehityksessä on monimutkainen. Granulosa-soluilla on androgeenireseptoreita. Nämä eivät ole ainoastaan substraatteja FSH:n indusoimalle estrogeenien aromatisaatiolle, vaan ne voivat myös tehostaa aromatisaatiota pieninä pitoisuuksina. Kun androgeenitasot nousevat, preantraaliset granulosa-solut valitsevat ensisijaisesti aromatisaatioreitin sijaan estrogeenit, vaan yksinkertaisemman androgeenireitin 5a-reduktaasin kautta muuntaakseen ne androgeeniksi, jota ei voida muuntaa estrogeeniksi, mikä estää aromataasiaktiivisuutta. Tämä prosessi estää myös FSH- ja LH-reseptorien muodostumista, mikä pysäyttää follikkelien kehityksen.
Aromatisaatioprosessissa korkean androgeenipitoisuuden omaava follikkeli käy läpi atresiaprosesseja. Follikkelin kasvu ja kehitys riippuvat sen kyvystä muuntaa androgeeneja estrogeeneiksi.
FSH:n läsnä ollessa follikkelinesteessä vallitseva aine on estrogeenit. FSH:n puuttuessa androgeenit. LH:ta ei normaalisti ole follikkelinesteessä kierron puoliväliin asti. Heti kun LH-taso nousee, granulosasolujen mitoottinen aktiivisuus vähenee, ilmenee degeneratiivisia muutoksia ja follikkelin androgeenitaso nousee. Follikulaarinesteen steroiditaso on korkeampi kuin plasmassa ja heijastaa munasarjasolujen: granulosa- ja theca-solujen, toiminnallista aktiivisuutta. Jos FSH:n ainoa kohde on granulosasolut, LH:lla on monia kohteita - theca-solut, strooma- ja luteiinisolut sekä granulosasolut. Sekä granulosa- että theca-soluilla on kyky steroidogeneesiin, mutta aromataasiaktiivisuus on vallitsevaa granulosasoluissa.
Vasteena LH:lle theca-solut tuottavat androgeeneja, jotka sitten granulosa-solut muuttavat estrogeeneiksi FSH:n indusoiman aromatisaation kautta.
Follikkelin kasvaessa theca-solut alkavat ilmentää LH-reseptorien, P450 sec:n ja 3beta-hydroksisteroididehydrogenaasin geenejä. Insuliinimäinen kasvutekijä (IGF-1) lisää synergistisesti LH:n kanssa geenien ilmentymistä, mutta ei stimuloi steroidogeneesiä.
Munasarjojen steroidogeneesi on aina LH-riippuvaista. Follikkelin kasvaessa theca-solut ilmentävät P450c17-entsyymiä, joka muodostaa androgeenia kolesterolista. Granulosa-soluilla ei ole tätä entsyymiä, ja ne ovat riippuvaisia theca-soluista tuottaakseen estrogeenia androgeeneistä. Toisin kuin steroidogeneesi, follikulogeneesi on FSH-riippuvaista. Follikkelin kasvaessa ja estrogeenitasojen noustessa aktivoituu takaisinkytkentämekanismi - FSH:n tuotanto estyy, mikä puolestaan johtaa follikkelin aromataasiaktiivisuuden vähenemiseen ja lopulta follikkeliatresiaan apoptoosin (ohjelmoidun solukuoleman) kautta.
Estrogeenien ja FSH:n takaisinkytkentämekanismi estää jo kasvamaan alkaneiden follikkelien kehitystä, mutta ei dominoivan follikkelin kehitystä. Dominoiva follikkeli sisältää enemmän FSH-reseptoreita, jotka tukevat granulosasolujen lisääntymistä ja androgeenien aromatisoitumista estrogeeneiksi. Lisäksi parakriiniset ja autokriiniset reitit toimivat tärkeänä antraalisten follikkelien kehityksen koordinoijana.
Autokriininen/parakriininen säätelijä koostuu peptideistä (inhibiini, aktiini, follistatiini), joita granulosasolut syntetisoivat vasteena FSH:lle ja jotka pääsevät follikkelinesteeseen. Inhibiini vähentää FSH:n eritystä; aktiini stimuloi FSH:n vapautumista aivolisäkkeestä ja tehostaa FSH:n vaikutusta munasarjassa; follistatiini estää FSH:n aktiivisuutta, mahdollisesti sitoutumalla aktiiniin. Ovulaation ja keltarauhasen kehittymisen jälkeen inhibiini on LH:n säätelemässä.
Munasarjasolujen kasvuun ja erilaistumiseen vaikuttavat insuliinin kaltaiset kasvutekijät (IGE). IGF-1 vaikuttaa granulosasoluihin aiheuttaen syklisen adenosiinimonofosfaatin (cAMP), progesteronin, oksitosiinin, proteoglykaanin ja inhibiinin lisääntymistä.
IGF-1 vaikuttaa theca-soluihin aiheuttaen lisääntynyttä androgeenituotantoa. Theca-solut puolestaan tuottavat tuumorinekroositekijää (TNF) ja epidermaalista kasvutekijää (EGF), joita FSH myös säätelee.
EGF stimuloi granulosasolujen lisääntymistä. IGF-2 on follikkelinesteessä tärkein kasvutekijä, ja siinä on myös IGF-1:tä, TNF-a:aa, TNF-3:aa ja EGF:ää.
Munasarjojen toiminnan parakriinisen ja/tai autokriinisen säätelyn häiriö näyttää vaikuttavan ovulaatiohäiriöihin ja monirakkulaisten munasarjojen kehittymiseen.
Antraalisen follikkelin kasvaessa estrogeenipitoisuus follikkelinesteessä kasvaa. Niiden lisääntymisen huipulla LH-reseptoreita ilmestyy granulosasoluihin, granulosasolut luteinisoituvat ja progesteronin tuotanto lisääntyy. Niinpä preovulaatiovaiheessa estrogeenituotannon lisääntyminen aiheuttaa LH-reseptorien ilmaantumisen, LH puolestaan aiheuttaa granulosasolujen luteinisaatiota ja progesteronin tuotantoa. Progesteronin lisääntyminen vähentää estrogeenitasoja, mikä ilmeisesti aiheuttaa toisen FSH-piikin syklin puolivälissä.
Ovulaation uskotaan tapahtuvan 10–12 tuntia LH-huipun jälkeen ja 24–36 tuntia estradiolihuipun jälkeen. LH:n uskotaan stimuloivan munasolun jakautumista, granulosasolujen luteinisaatiota sekä progesteronin ja prostaglandiinin synteesiä follikkeleissa.
Progesteroni tehostaa proteolyyttisten entsyymien aktiivisuutta, jotka yhdessä prostaglandiinin kanssa osallistuvat follikkeliseinämän repeämiseen. Progesteronin indusoima FSH-piikki mahdollistaa munasolun poistumisen follikkelista muuttamalla plasminogeenin proteolyyttiseksi entsyymiksi plasmiiniksi ja tarjoaa riittävän määrän LH-reseptoreita luteaalivaiheen normaalia kehitystä varten.
Kolmen päivän kuluessa ovulaatiosta granulosa-solujen koko kasvaa, ja niihin ilmestyy tyypillisiä pigmentillä, luteiinilla, täytettyjä vakuoleja. Theca-luteal-solut erilaistuvat thekasta ja stroomasta ja niistä tulee osa keltarauhasta. Keltarauhaseen tunkeutuvat kapillaarit kehittyvät hyvin nopeasti angiogeneesitekijöiden vaikutuksesta, ja parantuneen verisuonituksen myötä progesteronin ja estrogeenien tuotanto lisääntyy. Steroidogeneesin aktiivisuus ja keltarauhasen elinikä määräytyvät LH-tason mukaan. Keltarauhanen ei ole homogeeninen solumuodostelma. Kahden keltarauhassolutyypin lisäksi se sisältää endoteelisoluja, makrofageja, fibroblasteja jne. Suuret keltarauhassolut tuottavat peptidejä (relaksiini, oksitosiini) ja ovat aktiivisempia steroidogeneesissä, ja niillä on suurempi aromataasiaktiivisuus ja suurempi progesteronin synteesi kuin pienillä soluilla.
Progesteronin huippu saavutetaan 8. päivänä LH-huipun jälkeen. On huomattava, että progesteronia ja estradiolia erittyy jaksottaisesti luteaalivaiheessa korreloinnissa LH:n pulssimaisen erityksen kanssa. Keltarauhasen muodostuessa inhibiinin tuotannon säätely siirtyy FSH:lta LH:lle. Inhibiini lisääntyy estradiolin lisääntyessä ennen LH-huippua ja jatkaa lisääntymistään LH-huipun jälkeen, vaikka estrogeenitasot laskevat. Vaikka granulosasolut erittävät inhibiiniä ja estradiolia, niitä säätelevät eri reitit. Inhibiinin lasku luteaalivaiheen lopussa edistää FSH:n lisääntymistä seuraavassa syklissä.
Keltarauhanen pienenee hyvin nopeasti - 9.-11. päivänä ovulaation jälkeen.
Rappeutumisen mekanismi on epäselvä eikä se liity estrogeenien luteolyyttiseen rooliin tai reseptoriin liittyvään mekanismiin, kuten kohdun limakalvolla on havaittu. Keltarauhasen tuottamien estrogeenien roolille on toinenkin selitys. Tiedetään, että estrogeenejä tarvitaan progesteronireseptorien synteesiin kohdun limakalvolla. Luteaalivaiheen estrogeenit ovat todennäköisesti välttämättömiä progesteroniin liittyville muutoksille kohdun limakalvolla ovulaation jälkeen. Progesteronireseptorien riittämätön kehitys riittämättömien estrogeenitasojen seurauksena on mahdollisesti lisämekanismi hedelmättömyydelle ja varhaisen raskauden keskenmenoille, jotka ovat toinen luteaalivaiheen puutoksen muoto. Uskotaan, että keltarauhasen elinikä vakiintuu ovulaation aikaan. Ja se varmasti taantuu, jos sitä ei tueta istukkahormonilla raskauden vuoksi. Näin ollen keltarauhasen regressio johtaa estradiolin, progesteronin ja inhibiinin pitoisuuksien laskuun. Inhibiinin lasku poistaa sen suppressoivan vaikutuksen FSH:hon; Estradiolin ja progesteronin lasku mahdollistaa GnRH-erityksen erittäin nopean palautumisen ja aivolisäkkeen takaisinkytkentämekanismin poistumisen. Inhibiinin ja estradiolin lasku yhdessä GnRH-pitoisuuden nousun kanssa lisää FSH:n tuotantoa LH:n sijaan. FSH:n nousu johtaa follikkelien kasvuun, minkä jälkeen valitaan dominoiva follikkeli, ja uusi sykli alkaa, jos raskautta ei synny. Steroidihormoneilla on keskeinen rooli lisääntymisbiologiassa ja yleisessä fysiologiassa. Ne määräävät ihmisen fenotyypin, vaikuttavat sydän- ja verisuonijärjestelmään, luuston aineenvaihduntaan, ihoon ja yleiseen hyvinvointiin ja niillä on keskeinen rooli raskaudessa. Steroidihormonien vaikutus heijastaa solunsisäisiä ja geneettisiä mekanismeja, jotka ovat välttämättömiä solunulkoisen signaalin välittämiseksi solun tumaan fysiologisen vasteen aikaansaamiseksi.
Estrogeenit tunkeutuvat diffuusisti solukalvon läpi ja sitoutuvat solun tumassa sijaitseviin reseptoreihin. Reseptori-steroidikompleksi sitoutuu sitten DNA:han. Kohdesoluissa nämä vuorovaikutukset johtavat geenien ilmentymiseen, proteiinisynteesiin ja tiettyihin solu- ja kudostoimintoihin.