Lääketieteen asiantuntija

Uudet julkaisut
Keuhkoputket
Viimeksi tarkistettu: 04.07.2025

Kaikki iLive-sisältö tarkistetaan lääketieteellisesti tai se tarkistetaan tosiasiallisen tarkkuuden varmistamiseksi.
Meillä on tiukat hankintaohjeet ja vain linkki hyvämaineisiin mediasivustoihin, akateemisiin tutkimuslaitoksiin ja mahdollisuuksien mukaan lääketieteellisesti vertaisarvioituihin tutkimuksiin. Huomaa, että suluissa ([1], [2] jne.) Olevat numerot ovat napsautettavia linkkejä näihin tutkimuksiin.
Jos sinusta tuntuu, että jokin sisältö on virheellinen, vanhentunut tai muuten kyseenalainen, valitse se ja paina Ctrl + Enter.
Oikea pääkeuhkoputki on eräänlainen henkitorven jatke. Sen pituus on 28–32 mm, ontelon halkaisija 12–16 mm. Vasen pääkeuhkoputki on 40–50 mm pitkä ja leveys 10–13 mm.
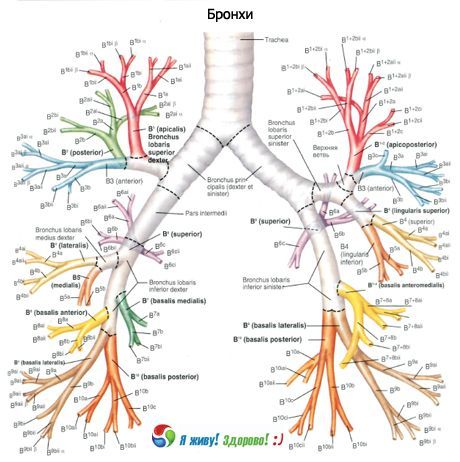
Perifeeriä kohti pääkeuhkoputket jakautuvat kaksijakoisesti lohko-, segmentti- ja subsegmenttiputkiin sekä edelleen terminaalisiin ja hengityskeuhkoputkiin. Kuitenkin esiintyy myös jakautumista kolmeen haaraan (trifurkaatio) ja useampaan.
Oikea pääkeuhkoputki jaetaan ylempään lohkoon ja välilohkoon, ja välilohko keskimmäiseen lohkoon ja alempaan lohkoon. Vasen pääkeuhkoputki jaetaan ylempään lohkoon ja alempaan lohkoon. Hengitysteiden sukupolvien kokonaismäärä vaihtelee. Pääkeuhkoputkesta alkaen ja keuhkorakkulapusseihin päättyen sukupolvien enimmäismäärä on 23–26.
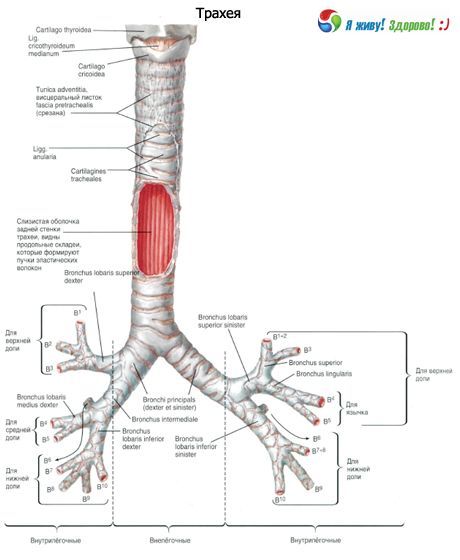
Pääkeuhkoputket ovat ensimmäisen asteen keuhkoputkia, lobar-keuhkoputket ovat toisen asteen keuhkoputkia, segmentaaliset keuhkoputket ovat kolmannen asteen keuhkoputkia jne.
4.–13. sukupolven keuhkoputkien halkaisija on noin 2 mm, ja niitä on yhteensä 400. Pääteilmanjohtimissa halkaisija vaihtelee 0,5–0,6 mm:n välillä. Hengitysteiden pituus kurkunpäästä acinuksiin on 23–38 cm.
Oikea ja vasen pääkeuhkoputki (bronchi principles dexter et sinister) alkavat henkitorven haarautumasta viidennen rintanikaman yläreunan tasolta ja ulottuvat vastaavasti oikean ja vasemman keuhkon hilumiin. Keuhkojen hilumin alueella kumpikin pääkeuhkoputki jakautuu lohkokeuhkoon (toisen asteen keuhkoputki). Aortankaari sijaitsee vasemman pääkeuhkoputken yläpuolella ja poskiontelon laskimo oikean yläpuolella. Oikea pääkeuhkoputki on pystysuorempi ja lyhyempi (noin 3 cm) kuin vasen pääkeuhkoputki (4-5 cm pitkä). Oikea pääkeuhkoputki on leveämpi (halkaisija 1,6 cm) kuin vasen (1,3 cm). Pääkeuhkoputkien seinämillä on sama rakenne kuin henkitorven seinämillä. Sisäpuolelta pääkeuhkoputkien seinämät ovat vuorattu limakalvolla ja ulkopuolelta peittyneet adventitialla. Seinien perustana ovat takaa sulkemattomat rustot. Oikeassa pääkeuhkoputkessa on 6-8 rustopuolirengasta, vasemmassa - 9-12 rustoa.
Henkitorven ja tärkeimpien keuhkoputkien hermotus: oikean ja vasemman toistuvan kurkunpään hermojen haarat ja sympaattiset rungot.
Verenkierto: alemman kilpirauhasvaltimon haarat, sisäinen rintavaltimo, rinta-aortta. Laskimoiden ulosvirtaus tapahtuu brakiokefaalisiin laskimoihin.
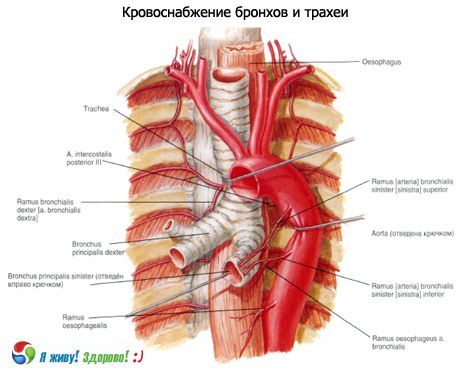
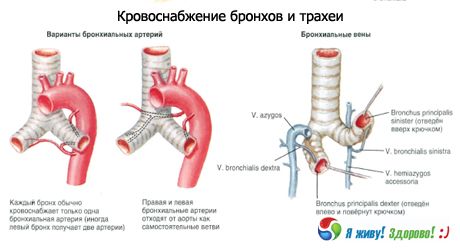
Imunestekierto: syviin kaulan lateraalisiin (sisäisiin jugulaarisiin) imusolmukkeisiin, pre- ja paratrakeaalisiin, ylempiin ja alempiin trakeobronkiaalisiin imusolmukkeisiin.
 [
[ Keuhkoputkien histologinen rakenne
Henkitorvea ja suuria keuhkoputkia peittää ulkopuolelta löysä sidekudostuppi, adventitia. Ulkokuori (adventitia) koostuu löysästä sidekudoksesta, joka sisältää suurten keuhkoputkien rasvasoluja. Sen läpi kulkevat imusuonet ja hermot. Adventitia ei ole selvästi rajautunut keuhkoputkien ympärillä olevaan sidekudokseen ja yhdessä keuhkoputkien kanssa mahdollistaa keuhkoputkien jonkinasteisen siirtymisen suhteessa ympäröiviin keuhkojen osiin.
Sisämpänä ovat sidekudos-rustoinen ja osittain lihaksikas kerros, limakalvonalainen kerros ja limakalvo. Kuitukerroksessa on rustomaisten puolirenkaiden lisäksi elastisten kuitujen verkosto. Henkitorven sidekudos-sidekudos on yhteydessä viereisiin elimiin löysän sidekudoksen avulla.
Henkitorven ja suurten keuhkoputkien etu- ja sivuseinät muodostuvat rustosta ja niiden välissä olevista rengasmaisista nivelsiteistä. Pääkeuhkoputkien rustomainen luuranko koostuu hyaliiniruston puolirenkaista, jotka pienenevät keuhkoputkien halkaisijan pienentyessä ja muuttuvat elastisiksi rustoiksi. Näin ollen vain suuret ja keskikokoiset keuhkoputket koostuvat hyaliinirustosta. Rusto vie 2/3 ympärysmitasta, kalvomainen osa 1/3. Ne muodostavat fibrokartilaginousisen luurangon, joka varmistaa henkitorven ja keuhkoputkien ontelon säilymisen.
Lihasniput keskittyvät henkitorven kalvomaiseen osaan ja pääkeuhkoputkiin. On erotettava pinnallinen eli ulompi kerros, joka koostuu harvinaisista pitkittäissuunnasta, ja syvä eli sisempi kerros, joka on poikittaisten kuitujen muodostama yhtenäinen ohut kalvo. Lihaskuidut eivät sijaitse ainoastaan ruston päiden välissä, vaan ne ulottuvat myös henkitorven rusto-osan ja suuremmassa määrin pääkeuhkoputkien rengasmaisiin tiloihin. Näin ollen henkitorvessa sileän lihaksen kimppuja, joilla on poikittainen ja vino järjestely, löytyy vain kalvomaisesta osasta, ts. lihaskerros sellaisenaan puuttuu. Pääkeuhkoputkissa on harvinaisia sileiden lihasten ryhmiä koko kehän ympärillä.
Keuhkoputkien halkaisijan pienentyessä lihaskerros kehittyy ja sen kuidut kulkevat hieman vinoon suuntaan. Lihasten supistuminen aiheuttaa paitsi keuhkoputkien luumenin kaventumisen, myös niiden lyhenemisen, minkä seurauksena keuhkoputket osallistuvat uloshengitykseen hengitysteiden kapasiteetin pienentymisen vuoksi. Lihasten supistuminen mahdollistaa keuhkoputkien luumenin kaventumisen 1/4:llä. Sisäänhengitettäessä keuhkoputki pitenee ja laajenee. Lihakset saavuttavat toisen asteen hengitysteiden keuhkoputket.
Lihaskerroksen sisällä on submukoosinen kerros, joka koostuu löysästä sidekudoksesta. Se sisältää verisuoni- ja hermomuodostelmia, submukoosisen imusuonten verkoston, imukudosta ja merkittävän osan keuhkoputkien rauhasista, jotka ovat tubulaari-asinoosista tyyppiä ja joilla on sekalaista limainen-seroottinen erite. Ne koostuvat pääteosista ja erityskanavista, jotka avautuvat pullonmuotoisiksi laajennuksiksi limakalvon pinnalla. Kanavien suhteellisen suuri pituus myötävaikuttaa keuhkoputkentulehduksen pitkäaikaiseen kulkuun rauhasten tulehdusprosessien aikana. Rauhasten surkastuminen voi johtaa limakalvon kuivumiseen ja tulehduksellisiin muutoksiin.
Eniten suuria rauhasia sijaitsee henkitorven haarautumiskohdan yläpuolella ja alueella, jossa pääkeuhkoputket jakautuvat lohkokeuhkoputkiksi. Terve ihminen erittää jopa 100 ml eritettä päivässä. Se koostuu 95 % vedestä ja 5 % on yhtä paljon proteiineja, suoloja, lipidejä ja epäorgaanisia aineita. Eritteessä on vallitsevasti musiineja (suurimolekyylisiä glykoproteiineja). Tällä hetkellä glykoproteiineja on 14 tyyppiä, joista 8 löytyy hengityselimistä.
Keuhkoputkien limakalvo
Limakalvo koostuu limakalvon epiteelistä, tyvikalvosta, limakalvon lamina propriasta ja lihaskalvosta.
Keuhkoputken epiteeli sisältää ylä- ja alasoluja, jotka ovat kiinnittyneet tyvikalvoon. Tyvikalvon paksuus vaihtelee 3,7:stä 10,6 μm:iin. Henkitorven ja suurten keuhkoputkien epiteeli on monirivinen, sylinterimäinen ja värekarvallinen. Epiteelin paksuus segmentaalisten keuhkoputkien tasolla vaihtelee 37:stä 47 μm:iin. Se koostuu neljästä pääsolutyypistä: värekarvallisista, pikarisoluista, välisoluista ja tyvisoluista. Lisäksi esiintyy seroosi-, harja-, Clara- ja Kulchitsky-soluja.
Värekarvalliset solut ovat vallitsevia epiteelikerroksen vapaalla pinnalla (Romanova LK, 1984). Niillä on epäsäännöllinen prisman muoto ja soikea, kuplan muotoinen tuma, joka sijaitsee solun keskiosassa. Sytoplasman elektronioptinen tiheys on pieni. Mitokondrioita on vähän, endoplasminen rakeinen retikulum on huonosti kehittynyt. Jokaisella solulla on lyhyitä mikrovilluksia ja noin 200 sädekarvallista siliaa pinnallaan, jotka ovat 0,3 μm paksuja ja noin 6 μm pitkiä. Ihmisellä silioiden tiheys on 6 μm 2.
Vierekkäisten solujen väliin muodostuu tyhjiä tiloja; solut ovat yhteydessä toisiinsa sytoplasman ja desmosomien sormimaisten ulokkeiden avulla.
Silmäsolujen populaatio jaetaan seuraaviin ryhmiin niiden apikaalisen pinnan erilaistumisasteen mukaan:
- Solut tyvikappaleiden ja aksoneemien muodostumisvaiheessa. Tällöin apikaalisella pinnalla ei ole enää silioita. Tänä aikana tapahtuu sentriolien kertymistä, jotka siirtyvät solujen apikaaliselle pinnalle ja muodostuvat tyvikappaleet, joista silioiden aksoneemit alkavat muodostua.
- Solut kohtalaisen voimakkaan siliogeneesin ja silioiden kasvun vaiheessa. Tällaisten solujen apikaaliselle pinnalle ilmestyy pieni määrä silioita, joiden pituus on 1/2-2/3 erilaistuneiden solujen silioiden pituudesta. Tässä vaiheessa mikrovillukset ovat vallitsevia apikaalisella pinnalla.
- Solut aktiivisen silioiden muodostumisen ja silioiden kasvun vaiheessa. Tällaisten solujen apikaalinen pinta on lähes kokonaan silioiden peitossa, joiden koot vastaavat solujen silioiden kokoja edellisessä silioiden muodostumisvaiheessa.
- Solut, jotka ovat täydellisen silioiden muodostumisen ja silioiden kasvun vaiheessa. Tällaisten solujen apikaalinen pinta on kokonaan peittynyt tiheästi järjestetyillä pitkillä silioilla. Elektronidiffraktiokuviot osoittavat, että vierekkäisten solujen siliat ovat suuntautuneet samaan suuntaan ja kaarevat. Tämä on osoitus limakalvojen kuljetuksesta.
Kaikki nämä soluryhmät näkyvät selvästi valoelektronimikroskopialla (SEM) saaduissa valokuvissa.
Siliat ovat kiinnittyneet solun apikaalisessa osassa sijaitseviin tyvikappaleisiin. Silioiden aksoneema muodostuu mikrotubuluksista, joista 9 paria (dubletteja) sijaitsee reunoilla ja 2 yksittäistä (singlettejä) keskellä. Dubletit ja singletit ovat yhteydessä toisiinsa neksiinifibrillien avulla. Kummassakin dubletissa on toisella puolella kaksi lyhyttä "kahvaa", jotka sisältävät ATPaasia, joka osallistuu ATP-energian vapautumiseen. Tämän rakenteen ansiosta siliat värähtelevät rytmisesti 16-17 sekunnin taajuudella nenänielun suuntaan.
Ne liikuttavat epiteeliä peittävää limakalvoa noin 6 mm/min nopeudella varmistaen siten keuhkoputken jatkuvan tyhjennystoiminnan.
Useimpien tutkijoiden mukaan värekarvalliset epiteelisolut ovat lopullisen erilaistumisen vaiheessa eivätkä kykene jakautumaan mitoosin avulla. Nykyaikaisen käsitteen mukaan tyvisolut ovat välisolujen esiasteita, jotka voivat erilaistua värekarvallisiksi soluiksi.
Pikarisolut, kuten värekarvalliset solut, ulottuvat epiteelikerroksen vapaalle pinnalle. Henkitorven ja suurten keuhkoputkien kalvomaisessa osassa värekarvalliset solut muodostavat jopa 70–80 %, kun taas pikarisolujen osuus on enintään 20–30 %. Paikoissa, joissa henkitorven ja keuhkoputkien reunoilla on rustoisia puolirenkaita, löytyy vyöhykkeitä, joissa värekarvallisten ja pikarisolujen suhteet vaihtelevat:
- jossa on pääasiassa sädekehäisiä soluja;
- lähes yhtä suurella suhdeluvulla värekarvallisia ja erittäviä soluja;
- jossa on hallitseva määrä erittäviä soluja;
- jossa on täydellinen tai lähes täydellinen värekarvallisten solujen puuttuminen ("ei-värekarvaiset").
Pikarisolut ovat merokriinistä tyyppiä olevia yksisoluisia rauhasia, jotka erittävät limaa. Solun muoto ja tuman sijainti riippuvat eritysvaiheesta ja supranukleaarisen osan täyttymisestä liman rakeilla, jotka sulautuvat suuremmiksi rakeiksi ja joille on ominaista alhainen elektronitiheys. Pikarisoluilla on pitkänomainen muoto, joka eritteen kertymisen aikana muodostaa pikarin, jonka pohja sijaitsee tyvikalvolla ja on läheisesti yhteydessä siihen. Solun leveä pää työntyy kupolin muotoon vapaalle pinnalle ja on varustettu mikrovilluksilla. Sytoplasma on elektronitiheä, tuma on pyöreä ja endoplasminen retikulum on karkeaa ja hyvin kehittynyt.
Pikarisolut ovat jakautuneet epätasaisesti. Pyyhkäisyelektronimikroskopia paljasti, että epiteelikerroksen eri vyöhykkeillä on heterogeenisiä alueita, jotka koostuvat joko vain värekarvallisista epiteelisoluista tai vain erittävien solujen muodostamista soluista. Pikarisolujen jatkuvia kertymiä on kuitenkin suhteellisen vähän. Terveen ihmisen segmentaalisen keuhkoputken reunoilla on alueita, joilla värekarvallisten epiteelisolujen ja pikarisolujen suhde on 4:1–7:1, kun taas muilla alueilla tämä suhde on 1:1.
Pikarisolujen määrä vähenee distaalisesti keuhkoputkissa. Keuhkoputkissa pikarisolut korvautuvat Clara-soluilla, jotka osallistuvat liman ja alveolaarisen hypofaasin seroosien komponenttien tuotantoon.
Pienissä keuhkoputkissa ja keuhkoputkissa pikarisolut puuttuvat normaalisti, mutta niitä voi esiintyä patologiassa.
Vuonna 1986 tšekkiläiset tiedemiehet tutkivat kanien hengitysteiden epiteelin reaktiota erilaisten limaa irrottavien aineiden suun kautta antoon. Kävi ilmi, että limaa irrottavien aineiden kohdesolut ovat pikarisoluja. Liman poistamisen jälkeen pikarisolut yleensä rappeutuvat ja poistuvat vähitellen epiteelistä. Pikarisolujen vaurioaste riippuu annetusta aineesta: lasolvanilla on suurin ärsyttävä vaikutus. Bronkolysiinin ja bromheksiinin annon jälkeen hengitysteiden epiteelissä tapahtuu massiivista uusien pikarisolujen erilaistumista, mikä johtaa pikarisolujen hyperplasiaan.
Tyvi- ja välisolut sijaitsevat syvällä epiteelikerroksessa eivätkä yllä vapaalle pinnalle. Nämä ovat vähiten erilaistuneita solumuotoja, minkä vuoksi fysiologinen uudistuminen tapahtuu pääasiassa. Välisolujen muoto on pitkänomainen, tyvisolut ovat epäsäännöllisen kuutiomaisia. Molemmilla on pyöreä, DNA-rikas tuma ja pieni määrä sytoplasmaa, jonka tiheys on suurempi tyvisoluissa.
Tyvisolut kykenevät synnyttämään sekä sädekehäisiä että pikarisoluja.
Erittyvät ja värekarvalliset solut yhdistetään nimellä "mukosiliaarinen laite".
Liman liikkumista keuhkojen hengitysteissä kutsutaan mukosiliaariseksi puhdistumaksi. MCC:n toiminnallinen tehokkuus riippuu värekarvallisen epiteelin värekarvojen liikkumistiheydestä ja synkroniteetista sekä, mikä erittäin tärkeää, liman ominaisuuksista ja reologisista ominaisuuksista eli pikarisolujen normaalista erityskapasiteetista.
Seroosisoluja on vähän, ne ulottuvat epiteelin vapaalle pinnalle ja niille on tunnusomaista pienet elektronitiheät proteiinierityksen jyväset. Sytoplasma on myös elektronitiheä. Mitokondriot ja karkea retikulum ovat hyvin kehittyneet. Tuma on pyöreä ja sijaitsee yleensä solun keskiosassa.
Erittäviä soluja eli Clara-soluja on eniten pienissä keuhkoputkissa ja bronkioleissa. Kuten seroosisolut, ne sisältävät pieniä elektronitiheitä jyväsiä, mutta niille on ominaista sytoplasman alhainen elektronitiheys ja sileän, endoplasmisen retikulumin hallitsevuus. Pyöreä tuma sijaitsee solun keskiosassa. Clara-solut osallistuvat fosfolipidien muodostumiseen ja mahdollisesti pinta-aktiivisen aineen tuotantoon. Lisääntyneen ärsytyksen olosuhteissa ne voivat ilmeisesti muuttua pikarisoluiksi.
Harjassoluilla on vapaalla pinnalla mikrovilluksia, mutta niillä ei ole silioita. Niiden sytoplasmassa on alhainen elektronitiheys ja tuma on soikea ja kuplan muotoinen. Ham A.:n ja Cormack D.:n (1982) käsikirjassa niitä pidetään pikarisoluina, jotka ovat erittäneet omaa eritettään. Niille annetaan monia toimintoja: imeytyminen, supistuvuus, eritys, kemoreseptori. Niitä on kuitenkin käytännössä tutkittu vähän ihmisen hengitysteissä.
Kulchitsky-soluja esiintyy koko keuhkoputkessa epiteelikerroksen pohjalla, ja ne eroavat tyvisoluista sytoplasman alhaisen elektronitiheyden ja pienten rakeiden läsnäolon perusteella, jotka näkyvät elektronimikroskoopilla ja hopeakyllästetyllä valomikroskoopilla. Ne luokitellaan APUD-järjestelmän neurosekretorisiin soluihin.
Epiteelin alla on tyvikalvo, joka koostuu kollageenista ja ei-kollageenisista glykoproteiineista; se tukee ja kiinnittyy epiteeliin, osallistuu aineenvaihduntaan ja immunologisiin reaktioihin. Tyvikalvon ja sen alla olevan sidekudoksen kunto määrää epiteelin rakenteen ja toiminnan. Lamina propria on löysä sidekudoskerros tyvikalvon ja lihaskerroksen välissä. Se sisältää fibroblasteja, kollageenia ja elastisia kuituja. Lamina propria sisältää veri- ja imusuonia. Kapillaarit ulottuvat tyvikalvoon, mutta eivät lävistä sitä.
Henkitorven ja keuhkoputkien limakalvolla, pääasiassa oikeassa kalvossa ja rauhasten lähellä, submukoossa, on jatkuvasti vapaita soluja, jotka voivat tunkeutua epiteelin läpi luumeniin. Näistä lymfosyytit ovat vallitsevia, plasmasolut, histiosyytit, syöttösolut (labrosyytit) sekä neutrofiiliset ja eosinofiiliset leukosyytit ovat harvinaisempia. Lymfoidisolujen jatkuvaa läsnäoloa keuhkoputkien limakalvolla kutsutaan erityistermillä "keuhkoputkiin liittyvä imukudos" (BALT), ja sitä pidetään immunologisena suojaavana reaktiona hengitysteihin ilman mukana tunkeutuville antigeeneille.